Расстояние между ламелями мм 70
Выберите категорию: Все Матрасы » Среднежесткие матрасы » Жесткие матрасы » Хиты продаж » Недорогие матрасы » Матрасы Аскона »» Матрасы Аскона «Balance» »» Матрасы Аскона «Expert» »» Матрасы Аскона «Promo» »» Матрасы Аскона «Serta Natural Start» »» Матрасы Аскона «Terapia New» » Матрасы Вегас (Vegas) »» Матрасы Вегас «Relax» »» Матрасы Вегас «Ultra» »» Матрасы Вегас «Смарт» («Smart») »» Матрасы Вегас «Хит» («Hit») »» Матрасы детские Вегас «Кидс» («Kids») » Матрасы Барро (Barro) »» Матрасы Барро «Эконом» »» Матрасы Барро «Заказ» »» Матрасы Барро «Комфорт» »» Матрасы Барро «Элит» »» Матрасы Барро «Элит Премиум» »» Матрасы Барро «Элит Престиж» »» Матрасы Барро «Идеальный комфорт» »» Матрасы Барро «Люкс» »» Матрасы Барро «Сыр» »» Матрасы Барро «Детская серия» » Матрасы Белабеддинг (Belabedding) »» Беспружинные матрасы Белабеддинг »» Пружинные матрасы Белабеддинг » Матрасы Кондор »» Матрасы Кондор Hard »» Матрасы Кондор Medio »» Матрасы Кондор Mini »» Матрасы Кондор Multi »» Матрасы Кондор Tango »» Матрасы Кондор Беспружинные » Матрасы Сиеста (Siesta) »» Пружинные матрасы Сиеста »» Беспружинные матрасы Сиеста »» Круглые матрасы Сиеста » Матрасы Цветные сны »» Матрасы Цветные сны «Беспружинные матрасы» »» Матрасы Цветные сны «Оптима» »» Матрасы Цветные сны «Релакс» »» Матрасы Цветные сны «Эконом» »» Матрасы Цветные сны «Элит» » Матрасы Эос »» Матрасы Эос «Аспект» »» Матрасы Эос «Идеал» »» Матрасы Эос «Лайт» »» Матрасы Эос «Премьер» »» Матрасы Эос «Элит» » Матрасы BelSon »» Матрасы BelSon Классик »» Матрасы BelSon Классик Идеал »» Матрасы BelSon Элит » Матрасы Sonit »» Матрасы Sonit Family »» Матрасы Sonit Free »» Матрасы Sonit Grand »» Матрасы Sonit Just »» Матрасы Sonit Layer »» Матрасы Sonit Next »» Матрасы Sonit IPS » Матрасы Фабрика сна »» Матрасы Фабрика сна «Топ» »» Матрасы Фабрика сна «Sweet» »» Матрасы Фабрика сна «Golg» »» Матрасы Фабрика сна «Latex» »» Матрасы Фабрика сна «Kinder» » Матрасы Территория сна »» Матрасы Территория сна Concept »» Матрасы Территория сна Romance »» Матрасы Территория сна Relax »» Матрасы Территория сна Optima » Детские матрасы » Пружинные матрасы » Беспружинные матрасы Кровати » Кровати Anrex (Анрэкс) » Кровати Black Red White BRW (БРВ) » Кровати Loft » Кровати ЗМФ (ZMF) » Кровати Мебель-Неман Наматрасники » Наматрасники Барро » Наматрасники защитные Вегас «Protect» » Наматрасники модифицирующие Вегас «Transform» » Наматрасники Фабрика сна Основания для матрасов » Основания для матрасов Вегас Мебель LOFT Подушки » Подушки Белабеддинг » Подушки Вегас
Название:
Артикул:
Текст:
Размер, см: Все32,3×29,642×82 (овал)42x8344x8845x3547x33,447×37,549x3049x3350x3350x3452x3055x11560x4060x4160x12060x14061x4062,5x3362x3462x4263x3365x12566x4266x4370x5070x9570x14070x18070x18670x19070x19570x19870x20080x18080x18680x19080x19580x19880x20085x19088x19088x19588x20090x18090x18690x19090x19590x19890x20090x200, 140x200100x180100x186100x190100x195100x198100x200110x186110x190110x195110x200120x180120x186120x190120x195120x198120x200130x186130x190130x195130x200140x186140x190140x195140x198140x200150x186150x190150x195150x198150x200158x190158x195158x198158x200160x186160x190160x195160x198160x200170x186170x190170x195170x200180x186180x190180x195180x198180x200180x210190x186190x190190x195190x200195x200200x24200x90200x186200x190200x195200x198200x200250x24диаметр 200диаметр 210диаметр 220диаметр 230
Производитель: ВсеAnrexBarroBelabeddingBRWKondorLoftrelaxSiestaSonitVegasАсконаБелСонВегасЗМФМебель-НеманТерритория снаФабрика снаЦветные сныЭос
Новинка: Вседанет
Спецпредложение: Вседанет
Результатов на странице: 5203550658095
Показать
Каркас для кровати расстояние между ламелями 2,5 см
Металлическое основание для матраса. Буковые гнутоклеенные ламели с расстоянием между ламелями 2,5 см.
Буковые гнутоклеенные ламели с расстоянием между ламелями 2,5 см.Каркас-кровати (ламели под матрас, ортопедические основания) на металлической раме
Ортопедический ламельный каркас (каркас-кровать, каркас под матрас, ортопедическое основание) – это гибкие буковые ламели, которые закреплены в металлической цельносвареной раме. Ламели по краям входят в пластиковые «кармашки», которые в свою очередь крепятся к раме.
Рама покрыта стандартной краской — порошковой эмалью черного цвета. Каркасы комплектуются ножками цилиндрической формы (для устойчивости). Пластиковые подошвы на ножках исключают царапины и повреждения пола, а также помогают установить каркас в ровном положении при недостатках неровной поверхности, т.к. регулируют высоту до 1,5 см. На внутренней стороне ламельного каркаса находятся так называемые «косынки» металлические для фиксации правильной формы каркас-кровати.
Высота ножек ламельного каркаса – 23 см.
Высота каркас-кровати в сборке– 27 см.
Допустимая нагрузка на каркас – до 130 кг (на одно спальное место)
Гарантия на ламельный каркас – 1 год.
Односпальные ламелевые каркас-кровати (ортопедические основания), шириной до 100 см, имеют поперечину, под которой находится дополнительная опора – 5-я ножка. Общее количество ножек в односпальном ортопедическом основании – 5 штук.
Двуспальные ламелевые каркас-кровати , шириной от 120 см, имеют внутреннюю продольную раму, под которой крепятся для прочности две дополнительные ножки. Общее количество ножек в ортопедическом основании – 6 штук.
Изготовление ортопедических каркас-кроватей на ламелях возможно с различной высотой ножек (цена на каркас при этом не меняется). Эластичные ламели изготовлены из буковой клееной фанеры. Ширина ламели – 5,5 см, толщина – 8 мм.
Металлическое основание для матраса. Буковые гнутоклеенные ламели с расстоянием между ламелями 2,5 см.Каркас-кровати (ламели под матрас, ортопедические основания) на металлической раме…
Каркасы кроватей с ламелями — Кровати & Каркасы / Come-For™
Здоровый сон – залог бодрого самочувствия на протяжении всего дня, энергии и способности активно добиваться поставленных целей.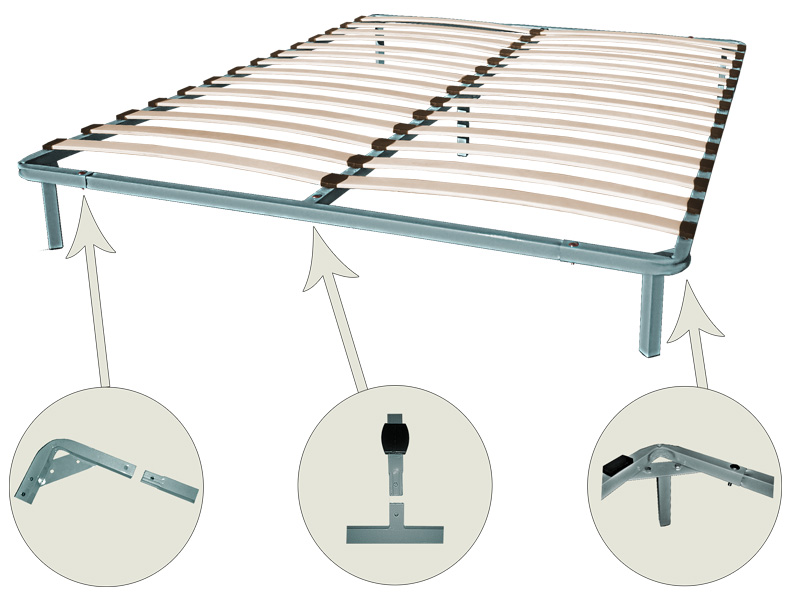 При покупке кровати нужно направлять внимание не только на ортопедические матрасы, но и на каркас кровати с ламелями. Правильно выбранная модель позволит забыть о вялом состоянии и сонливости по утрам, вы по-новому взглянете на окружающий вас мир. Каркас под кровать купить лучше всего у надежного изготовителя, который применяет качественные материалы и комплектующие.
При покупке кровати нужно направлять внимание не только на ортопедические матрасы, но и на каркас кровати с ламелями. Правильно выбранная модель позволит забыть о вялом состоянии и сонливости по утрам, вы по-новому взглянете на окружающий вас мир. Каркас под кровать купить лучше всего у надежного изготовителя, который применяет качественные материалы и комплектующие.
Купить каркас для кровати
Компания Come-for предлагает клиентам большой выбор ортопедических матрасов, наматрасников, одеял и подушек, каркасов кроватей. Каркасы для кровати с ламелями представлены от самых компактных размеров 80*190 см до просторных семейных 200*200 см. Приобретая каркас кровати, вы можете по достоинству оценить качество материла, соединительных элементов, что невозможно при покупке кровати или дивана в собранном виде.
Главное удобство состоит в том, что на одном сайте вы можете выбрать все, что вам нужно — каркас двуспальной кровати, ортопедический каркас для кровати и другие элементы. Come-for заботится о высоком уровне сервиса и комфорте покупателей, всегда идет навстречу их пожеланиям и предпочтениям. Доставка и оплата возможны разными способами, клиент сам выбирает для себя наиболее приемлемые.
Come-for заботится о высоком уровне сервиса и комфорте покупателей, всегда идет навстречу их пожеланиям и предпочтениям. Доставка и оплата возможны разными способами, клиент сам выбирает для себя наиболее приемлемые.
Особенности металлических каркасов с ламелями
Каркасы для кроватей с ламелями пользуются большой популярностью благодаря выраженному ортопедическому эффекту. Они способны повторять анатомическую структуру тела человека, что содействует спокойному сну, который полностью восстанавливает силы. При выборе такой кровати ориентируются на вес тела человека и особенности его телосложения. Такой индивидуальный подход позволяет в результате создать кровать, которая прослужит своему хозяину долгие годы и будет положительно влиять на здоровье позвоночника.
У каркасов может быть разное расстояние между ламелями, в зависимости от этого выбирается пружинный или беспружинный матрас. Когда приобретается каркас для кровати с ламелями, то стоит оценивать такие критерии:
— упругость, практичность и надежность ламелей;
— применение экологически чистых материалов и инновационных технологий;
— промежуток между ламелями;
— размер мебели, куда будет встраиваться каркас.
Металлические каркасы для кроватей с ламелями не требуют особенного ухода, их легко устанавливать и демонтировать, но при этом они демонстрируют значительный предел нагрузки.
ортопедические ламели матрасы ортопедические из латекса latex подушки каркас кровати :
Ортопедическое основание, что это?
Ортопедическая решетка (основание под матрас) представляет собой металлическую или деревянную раму с ножками или без ножек. Поперек рамы расположены рейки-ламели, на которых и лежит матрас.
Ламели могут быть пружинящими, из березы или бука, в этом случае ощущения от матраса становятся более «мягкими». Ламель может быть «узкой» (3-4 см) и «широкой» (7-9 см). Первая рекомендуется для матрасов с пружинами Multipocket; однако по опыту, и обычная «широкая» ламель прекрасно работает для всех типов матрасов.
Зачем нужны ортопедические основания?
Во-первых, ортопедические основания специально спроектированы для того, чтобы Вы максимально почувствовали ортопедический эффект матраса. Матрасы нельзя класть на кривую поверхность типа продавленных диванов, провисших кроватных сеток и дырявых поролоновых подушек. Во всех вышеперечисленных случаях спать на матрасе будет неудобно, он не даст ожидаемого ортопедического эффекта, плюс во время эксплуатации матраса очень быстро образуются вмятины и матрас потеряет ортопедические качества. Во-вторых, зачастую кровати как отечественного, так и импортного производства не комплектуются основанием, и его приходится докупать дополнительно. Поэтому при покупке кровати обязательно уточняйте у продавца, есть в конкретной модели лемели, или нет. В-третьих, ортопедический каркас обеспечивает свободную циркуляцию воздуха. Ведь за ночь человек выделяет 0,5-1 литр пота. Поэтому если матрас положить на основания без ламелей (на фанеру), то он будет хуже «дышать». В-четвертых, ортопедические основания позволяют сэкономить на кровати. На нем можно отлично спать, просто не будет декоративного изголовья, боковин и т.д. и т.п. В общем-то, кровать можно докупить и потом, при появлении средств, или когда вы определитесь с моделью.
Конструкционные различия ортопедических решеток:
|
 При выборе каркасов следует учесть, что основания из бука более красивы и оптимально подходят к ортопедическому матрасу, каркас из железа дешевле и немножко проигрывает во внешнем виде, но при этом не теряет своих ортопедических качеств.
При выборе каркасов следует учесть, что основания из бука более красивы и оптимально подходят к ортопедическому матрасу, каркас из железа дешевле и немножко проигрывает во внешнем виде, но при этом не теряет своих ортопедических качеств.
 Также имеет значение, сколько слоев материала используется для изготовления ламели. Самое оптимальное — это больше шести слоев. Ламели, сделанные из березы, являются более дешевыми, менее прочными, срок службы таких ламелей намного меньше.
Также имеет значение, сколько слоев материала используется для изготовления ламели. Самое оптимальное — это больше шести слоев. Ламели, сделанные из березы, являются более дешевыми, менее прочными, срок службы таких ламелей намного меньше.

Металлический каркас |
Деревянный каркас |
Тип ламелей
Прямые ламели |
Пружинные ламели шириной 7 см |
Узкие пружинные ламели шириной 3 см |
Крепление ламелей
Врезное крепление |
Накладное пластиковое крепление |
Накладное резиновое крепление |
Механизм трансформации
Без трансформации |
Ручной механизм трансформации |
Механизм трансформации с электроприводом |
Наличие ножек
Ортопедическое основание с ножками |
Ортопедическое основание без ножек |
Возможность изменения жёсткости ламелей
Без возможности изменения жёсткости ламелей |
С возможностью изменения жёсткости ламелей в отдельной части |
С возможностью изменения жёсткости ламелей по всей длине |
Анонсы статей
Боль в спине — как помочь себе.


Сложно сказать, кто чаще страдает от болезней спины — мужчины или женщины. У мужчин обострения чаще всего связаны с превышением допустимой нагрузки на позвоночник — поднял тяжелое на даче, потаскал на спине ребенка и т.д. Причины проблем со спиной у женщин чаще всего другие — повышенная постоянная нагрузка на спину в связи с работой по дому сопряжена с такими отягчающими обстоятельствами, как постоянное ношение обуви на каблуке, отказ от спорта (натренированные мышцы лучше поддерживают позвоночник), невнимание к своему здоровью. Так постепенно нашим постоянным спутником становится боль в спине. Если боли возникают незначительные и непродолжительные, мы чаще всего не обращаем на них должного внимания.
далее ››
Латексные матрасы: новый уровень комфорта.
Если мы говорим «качественный матраc» — подразумеваем «ортопедический матраc». Сегодня именно ортопедические матрацы пользуются наибольшим спросом, — они дарят нам полноценный отдых, «подстраиваясь» под особенности конституции и обеспечивая правильное положение нашего позвоночника во время сна.
Лидером материалов для изготовления ортопедических матрасов смело можно назвать латекс. Монолитный латексный блок является основой для беспружинных матраcов. А в пружинных матраcах с независимым блоком пружин латекс, наряду с другими материалами, выступает в качестве прослойки-наполнителя. Какими же свойствами обладает латекс? Что сулит потребителю наличие латексного наполнителя в матрасе?
Сегодня именно ортопедические матрацы пользуются наибольшим спросом, — они дарят нам полноценный отдых, «подстраиваясь» под особенности конституции и обеспечивая правильное положение нашего позвоночника во время сна.
Лидером материалов для изготовления ортопедических матрасов смело можно назвать латекс. Монолитный латексный блок является основой для беспружинных матраcов. А в пружинных матраcах с независимым блоком пружин латекс, наряду с другими материалами, выступает в качестве прослойки-наполнителя. Какими же свойствами обладает латекс? Что сулит потребителю наличие латексного наполнителя в матрасе?
далее ››
Каркас-кровати
Мы рады Вам сообщить, что по многочисленным просьбам, мы изменили модельный ряд наших каркасов.Так, теперь у нас каркасы не только из металлической трубы 25х25 мм, но и из трубы размером 30х30 мм.
Также добавилось количество ламелей с 22 до 24 на каркасах с минимальным расстоянием между ламелями, что обеспечивает зазор 25 мм.

Также рады сообщить вам о появлении у нас каркасов с длиной ламели 1000 мм. Появились на складе каркасы шириной 1000 мм и 2000 мм.
По умолчанию каркас 2000х2000 мы изготавливаем из двух частей, то есть разборной для удобства транспортировки.
Стандартное количество опор односпального каркаса -5 шт., двуспального — 6 шт. Однако На всех наших двуспальных каркасах имеется возможность установки двух дополнительных опор по длинной стороне периметра каркаса, то есть всего 8 шт. Опоры продаются поштучно.
Также сообщаем Вам о значительном снижении стоимости опор.
Также сообщаем вам, что можем изготавливать без удорожания нестандартные размеры каркасов, ближайшие к стандартным. Также за небольшую дополнительную оплату готовы добавить поперечные перемычки из металла.
Мы используем буковые ламели ( латофлекс ) 1 сорта. Толщина 8 мм, ширина 53 мм либо 68 мм. Соответственно и количество ламелей при одинаковых зазорах между ламелями может быть разным.
Например при длине каркаса 2000 мм:
— расстояние между ламелями в 2,5 см обеспечивает ламель шириной 53 мм — в количестве 24 шт. или ламель шириной 68 мм — в количестве 19 шт.;
— расстояние между ламелями в 4 см обеспечивает ламель шириной 53 мм — в количестве 19 шт. или ламель шириной 68 мм — в количестве 15 шт.;
— расстояние между ламелями в 6 см обеспечивает ламель шириной 53 мм — в количестве 15 шт. или ламель шириной 68 мм — в количестве 13 шт.;
Стандартная высота опоры 235 мм. Диаметр 50 мм. Используется регулируемый пластиковый подпятник с возможностью регулировки до 15 мм. Соответственно высота каркаса с ножками составляет 270-290 мм.
Мы можем без изменения стоимости изготовить для вас опоры до 300 мм.
Расстояние (зазор) между ламелями на наших каркас — кроватях мы предлагаем в 3-х вариантах:
6 см — отлично подойдет для матрацев с пружинами типа боннель, т.к диаметр такой пружины больше 6, 5 см;
4 см — универсальный размер, подходящий ко всем типам матрацев ;
2,5 см — этот размер для людей с весом от 100 кг или любителей запаса прочности.
Однако не надо забывать, что чем меньше расстояние между ламелями, тем каркас-кровать крепче, но ортопедичность меньше.
Выбор за вами.
РОЗНИЦА| 13.09.21 | Металл.труба 25х25 | Металл.труба 30х30 | |||
| РОЗНИЦА |
Между ламелями 6,5 см |
Между ламелями 4,5 см |
Между ламелями 2,5 см |
Между ламелями 4,5 см |
Между ламелями 2,5 см |
| Опоры продаются отдельно по цене 32 гривен за штуку | |||||
| Каркас 1900 х 800 | 853 | 902 | 965 | 969 | 1032 |
| Каркас 1900 х 900 | 879 | 923 | 998 | 995 | 1065 |
| Каркас 1900 х 1000 | 957 | 1019 | 1101 | 1088 | 1168 |
| Каркас 1900 х 1200 | 1434 | 1508 | 1604 | 1639 | 1734 |
| Каркас 1900 х 1400 | 1463 | 1541 | 1648 | 1672 | 1779 |
| Каркас 1900 х 1600 | 1493 | 1578 | 1696 | 1708 | 1827 |
| Каркас 1900 х 1800 | 1600 | 1700 | 1826 | 1830 | 1955 |
| Каркас 2000 х 800 | 861 | 909 | 975 | 976 | 1043 |
| Каркас 2000 х 900 | 891 | 938 | 1009 | 1006 | 1076 |
| Каркас 2000 х 1000 | 968 | 1039 | 1119 | 1106 | 1187 |
| Каркас 2000 х 1200 | 1449 | 1523 | 1615 | 1654 | 1745 |
| Каркас 2000 х 1400 | 1478 | 1559 | 1663 | 1690 | 1794 |
| Каркас 2000 х 1500 | 1511 | 1604 | 1715 | 1734 | 1845 |
| Каркас 2000 х 1600 | 1511 | 1604 | 1715 | 1734 | 1845 |
| Каркас 2000 х 1800 | 1618 | 1711 | 1847 | 1841 | 1978 |
| Каркас 2000 х 2000 | 1840 | 1973 | 2139 | 2103 | 2270 |
ОПТ
13. 09.21 09.21 |
Металл.труба 25х25 | Металл.труба 30х30 | |||
| ОПТ. |
Между ламелями 6,5 см |
Между ламелями 4,5 см |
Между ламелями 2,5 см |
Между ламелями 4,5 см |
Между ламелями 2,5 см |
| Опоры продаются отдельно по цене 30 гривен за штуку | |||||
| Каркас 1900 х 800 | 825 | 872 | 933 | 937 | 998 |
| Каркас 1900 х 900 | 850 | 893 | 965 | 962 | 1031 |
| Каркас 1900 х 1000 | 926 | 986 | 1065 | 1052 | 1130 |
| Каркас 1900 х 1200 | 1387 | 1459 | 1552 | 1585 | 1677 |
| Каркас 1900 х 1400 | 1415 | 1491 | 1595 | 1617 | 1721 |
| Каркас 1900 х 1600 | 1445 | 1526 | 1641 | 1652 | 1767 |
| Каркас 1900 х 1800 | 1548 | 1644 | 1766 | 1770 | 1891 |
| Каркас 2000 х 800 | 833 | 880 | 943 | 945 | 1009 |
| Каркас 2000 х 900 | 862 | 907 | 976 | 974 | 1041 |
| Каркас 2000 х 1000 | 936 | 1005 | 1082 | 1070 | 1149 |
| Каркас 2000 х 1200 | 1402 | 1474 | 1562 | 1600 | 1689 |
| Каркас 2000 х 1400 | 1430 | 1509 | 1609 | 1635 | 1735 |
| Каркас 2000 х 1500 | 1462 | 1552 | 1659 | 1677 | 1785 |
| Каркас 2000 х 1600 | 1462 | 1552 | 1659 | 1677 | 1785 |
| Каркас 2000 х 1800 | 1565 | 1656 | 1787 | 1781 | 1913 |
| Каркас 2000 х 2000 | 1780 | 1909 | 2070 | 2034 | 2196 |
EMM — Каркасы кроватей с ламелями
Каркасы кроватей с ламелями обладают исключительными амортизационными свойствами, гарантируют высокое качество сна и пользуются заслуженной популярностью. Они рекомендованы людям любого возраста и отлично сочетаются с ортопедическими матрасами всех типов.
Они рекомендованы людям любого возраста и отлично сочетаются с ортопедическими матрасами всех типов.
Конструктивные особенности
Металлический каркас с ламелями представляет собой ортопедическое основание спального места, функция которого – повышение комфортабельности кровати и увеличение срока службы матраса. Основа каркаса – прочная цельносварная или сборная рама, на которую параллельно друг другу крепятся гибкие ламели из фанеры, обеспечивающие необходимый ортопедический эффект.
Для изготовления ламелей применяется фанера из:
- бука – этот материал обладает повышенной прочностью и может выдерживать значительные нагрузки, но стоит дороже;
- березы – березовая фанера доступней по цене, отличается повышенной гибкостью и лучше амортизирует, но уступает буку по показателям прочности и долговечности.
Ламели могут быть разной ширины: широкие лучше «работают» в сочетании с беспружинными матрасами и моделями Боннель, узкие же рекомендованы для изделий с независимым блоком, у которых плотность пружин на квадратный метр более 500 шт. А лучший воздухообмен и, соответственно, гигиеничность спального места обеспечивают каркасы с ламелями, закрепленными поверх рамы (их можно заказать в нашем магазине с доставкой, а прайс на услугу есть на странице «Доставка и оплата»).
А лучший воздухообмен и, соответственно, гигиеничность спального места обеспечивают каркасы с ламелями, закрепленными поверх рамы (их можно заказать в нашем магазине с доставкой, а прайс на услугу есть на странице «Доставка и оплата»).
Актуальные варианты
Среди предлагаемых производителями каркасов под матрас наиболее востребованными являются:
- Бюджетные модели с ножками-опорами и без.
- Подъемные основания – они оснащаются газлифтами и актуальны для кроватей со встроенным бельевым коробом.
- Усиленные блоки с увеличенным количеством ламелей и уменьшенным расстоянием между ними – такие конструкции выдерживают высокие статические и динамические нагрузки.
- Изделия премиум-класса с двойными ламелями – они позволяют «настроить» жесткость каждой зоны спального места.
Каркасы кроватей: заказ онлайн с доставкой
На sleep-fly.com.ua представлены самые популярные модели каркасов кроватей с ламелями, которые можно заказать онлайн с доставкой до двери. А, чтобы получить дополнительную информацию по продукции, вы можете связаться с нами.
А, чтобы получить дополнительную информацию по продукции, вы можете связаться с нами.
Как выбрать правильный матрас
Сон, наравне с едой, водой и воздухом – базовая потребность каждого человека. Прерывающийся, некачественный сон пагубно влияет на физиологию и психику человека и может привести к необратимым изменениям в эндокринной системе. Со стороны психики некачественный сон приводит к развитию депрессивного и тревожно-фобического расстройств. Снижается качество жизни, человек становится более рассеянным и раздраженным. Доказано, что качество сна и правильный режим дня могут предупреждать развитие сахарного диабета.
Как выбрать матрас?
На качество сна влияет множество факторов, но люди часто забывают о том, на чем они спят. Казалось бы, какое значение имеет правильно подобранный матрас, если спать можно на любой более или менее подходящей поверхности?
Этот распространенный миф мешает людям найти именно то, что в разы улучшит не только их сон, но и положительно повлияет на здоровье в часы бодрствования.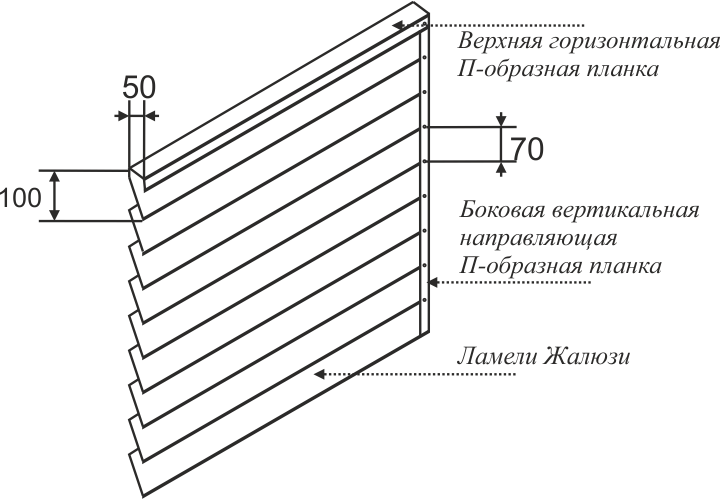 В этой статье мы докажем нужность матрасов с ортопедическим эффектом и поможем подобрать идеальные товары для сна, не переплачивая лишних денег.
В этой статье мы докажем нужность матрасов с ортопедическим эффектом и поможем подобрать идеальные товары для сна, не переплачивая лишних денег.
Кому нужен матрас
Вообще ортопедический матрас для ежедневного сна пригодится всем людям, независимо от медицинских показаний. Спать можно и на раскладушке, и на продавленном диване, но о комфорте, а тем более пользе, тут речи идти не может. Постоянная боль в плечах и пояснице, слабость и усталость после сна – верные признаки того, что вы спите на неправильной поверхности.
Внимательными нужно быть людям с хроническими заболеваниями опорно-двигательного аппарата: остеохондроз, сколиоз, межпозвонковая грыжа требуют к себе особого внимания. И купить ортопедический матрас – важная профилактическая мера, он сведет к минимуму неприятные проявления некоторых болезней позвоночника.
Какой матрас лучше выбрать?
Подбор матраса зависит от возраста человека и его веса. Ниже приведены несколько советов, которые гарантируют вам комфортный здоровый сон.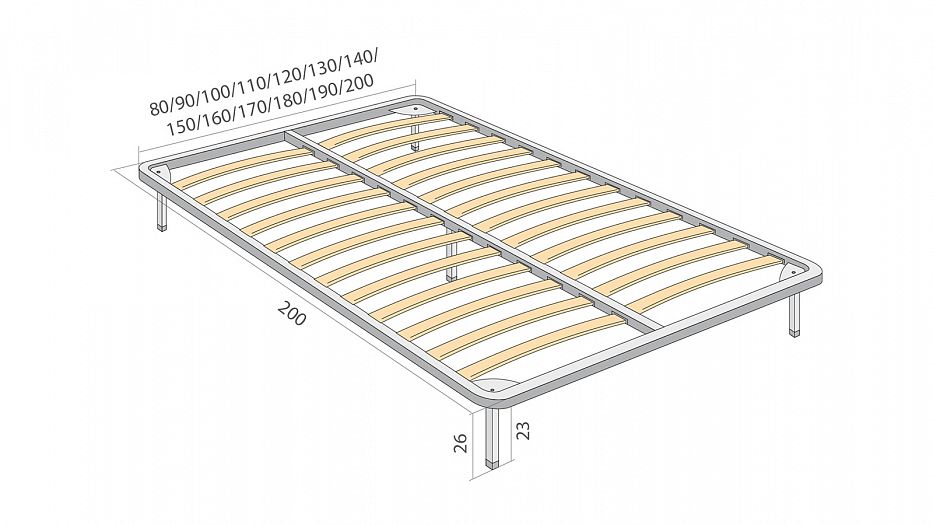
Перед покупкой матраса, обязательно полежите на нем. Странно читать такое в интернет-магазине, не так ли? Да, можно подобрать матрас по другим критериям, по которым матрасы подбирают консультанты интернет-магазина. Но тактильные ощущения очень важны, особенно когда речь идет об ортопедическом эффекте и степени жесткости матраса. Матрас должен идеально повторять силуэт тела, не оказывая обратного давления на мягкие ткани и одновременно гарантировать сильную поддержку позвоночнику, чтобы человек не проваливался в него как в гамак.
Как правильно выбрать матрас для кровати?
Немаловажно, чтобы матрас совпадал по размерам с кроватью. Если матрац больше на пару сантиметров, то возможна деформация мягких слоев матраса. Если у кровати имеется ортопедическое основание с ламелями, расстояние между ними должно быть не болеее 5 см, чтобы независимые пружины не проваливались в зазоры между ламелями.
Если у Вас есть возможность посетить салон хорошего сна «Сонит» — обязательно полежите на каждом из представленных на экспозиции матрасе.
Проверенная техника определения подходящей модели: если человек лежа на спине может свободно провести ладонью между матрасом и спиной в области талии — такой матрас через чур жесткий. Если вы утопаете в матрасе и на нем сложно лечь на бок – матрас слишком мягкий и надо выбрать другой.
Малейшие дискомфортные или болевые ощущения при «примерке» матраса говорит о том, что Вам необходимо подыскать другую модель.
Обратите внимание на чехол изделия и на наличие ручек. Чехол предпочтительно выбирать несъемный. Ручки нужны для переворачивания матраса. Каждые три месяца нужно менять стороны голова-ноги, а каждые пол года переворачивать, чтобы восстанавливать мягкие слои наполнения матраса.
Важны размеры и форма матраса. Покупайте матрас под размер спального места, чтобы он не деформировался в процессе эксплуатации.
У нас можно заказать матрасы нестандартных форм и размеров – для детских кроваток, матрас в автомобиль, круглые матрасы и др.
Из чего состоит матрас?
Важный фактор при выборе матраса для сна – наполнение изделия. Условно матрасы делятся на пружинные и беспружинные. Беспружинные представлены в виде монолита натурального латекса или анатомических пен. Пружинные блоки тоже различаются:
Условно матрасы делятся на пружинные и беспружинные. Беспружинные представлены в виде монолита натурального латекса или анатомических пен. Пружинные блоки тоже различаются:
- Бюджетный блок зависимых пружин типа Боннель.
- Блок независимых пружин TFK – каждая пружина в блоке зашита в отдельный чехол и при надавливании на одну пружину, ее соседки не продавливаются, что создает тот самый ортопедический эффект. Стандартный срок службы блока – 20 лет, но матрасы с ППУ служат не дольше 10 лет – они накапливают пыль и их все равно надо менять по мере необходимости.
- Блок независимых пружин TFK 7 зон комфорта (жесткости) – зонированный блок, пружины в нем имеют разный диаметр и толщину проволоки, что усиливает анатомические и ортопедические свойства матраса.
- Блок независимых пружин Multipocket точечно распределяет нагрузку на матрас за счет увеличенного количества пружин. Количество пружин может составлять 510 пружин/м² или 1000 пружин/м². Каждому участку тела уделяется в несколько раз больше внимания, что дает высокий ортопедический эффект, максимальную подстройку под каждый изгиб тела, снятие напряжения с позвоночника и полноценный сон.

Помимо пружинных блоков, в ортопедических матрасах присутствуют и другие наполнители, которые повышают комфортность матраса. Выделяют синтетические и натуральные наполнители.
Натуральные:
- Кокосовая койра – применяется для повышения жесткости матраса. Матрасы с койрой показаны людям со сколиозом и детям (для правильного формирования позвоночника). Обязательно поинтересуйтесь у консультантов, какой длины кокосовая койра – по ГОСТу волокно должно быть не короче 1 см. Кокосовая плита не должна напоминать опилки.
- Латекс – натуральный материал,проверенный годами и тысячами производителей товаров для сна. Ценится латекс за свои анатомические свойства и долговечность. Гипоалергенный, устойчивый к пылевому клещу, термостабильный материал. Благодаря перфорации отлично пропускает воздух и не накапливает лишнюю влагу. Выбирая латексный матрас, обратите внимание на содержание сока гевеи. Стопроцентным латексом считается материал, на 20 % состоящий из натурального сока гевеи. Срок службы монолитного латексного матраса составляет более 20 лет.
- Конский волос – используется как альтернатива кокосовой койры. Природный материал, но имеет ряд минусов – дорогой, плохо держит форму и может вылезать из чехла матраса. Пропылесосить такой матрас практически невозможно.
 Срок службы монолитного латексного матраса составляет более 20 лет.
Срок службы монолитного латексного матраса составляет более 20 лет.
Пенополиуретаны экстра-класса
Помимо натуральных материалов в производстве матрасов и диванов применяется такой наполнитель как анатомические пены. В своих анатомических и гипоалергенных свойствах они практически не отличаются от натурального латекса. Преимуществом пенополиуретанов — цена. Заказав матрас на основе анатомических пен, можно гарантировать себе здоровый сон, без лишней траты денег.
Каким должен быть матрас?
Выбор матраса — не мудрёная наука, и если знать основные критерии, то вы легко подберете нужную модель.
Матрас должен быть:
✔ Удобным и приятным на ощупь.
✔ Сделан из качественных, безопасных материалов.
✔ Долговечным.
✔ Эргономичным и простым в уходе.
✔ Эстетически привлекательным.
Если у вас остались вопросы – обязательно свяжитесь с консультантом интернет-магазина, который даст более подробную информацию о том, какой матрас подойдет вам.
Ламельные осветлители — обзор
18.2 Сточные воды пищевой промышленности и производства напитков
В пищевой промышленности и производстве напитков отработанные воды, образующиеся при различных операциях (переработка фруктов, очистка резервуаров и трубопроводов, а также мойка бутылок), смешиваются вместе, прежде чем они будут сбрасывается в городскую канализацию.
Эти сточные воды можно определить как высокопрочные сточные воды из-за их высоких значений ХПК и высокого содержания биоразлагаемых соединений, таких как азот или фосфорные элементы.Их можно сбрасывать в окружающую среду после специальной обработки для уменьшения количества загрязняющих веществ в соответствии со стандартами, установленными экологическими нормативами.
Использование MBR — многообещающая технология очистки высокопрочных сточных вод в качестве альтернативы традиционной обработке активным илом. Основные принципы, конфигурации и типичные применения систем MBR анализируются и обсуждаются в главе 6. Как подчеркивается в этой главе, наилучшая производительность MBR может быть достигнута за счет оптимизации рабочих параметров, таких как время гидравлического удерживания, время удерживания твердого вещества (SRT), смешанное щелочные взвешенные твердые частицы (MLSS) и трансмембранное давление (TMP).Системы предварительной обработки, физическая и химическая очистка и восстановители биологического обрастания, такие как порошкообразный активированный уголь, — все это системы, которые можно использовать для уменьшения явления загрязнения, тем самым повышая производительность MBR (Katayon et al., 2004; Mutamim, Noor, Abu Hassan, & Olsson, 2012).
Гибридный процесс производства питьевой воды из отработанных технологических вод предприятий по производству фруктовых соков был разработан Blöcher et al. (2002) в промышленном масштабе после предварительных исследований в пилотном масштабе (Noronha, Britz, Mavrov, Janke, & Chmiel, 2002).Отработанные воды малых и средних предприятий смешивались в уравнительном баке и очищались с помощью пластинчатого осветлителя для удаления большей части твердых частиц. Осветленные сточные воды обрабатывали в MBR, оборудованном погруженными трубчатыми мембранами MF (размер пор 0,04 мкм), в которых полностью задерживалась активная биомасса и другие твердые частицы. Хотя уровень содержания органических веществ в отработанной воде варьировался в широких пределах (от 1800 до 6600 мг / л), ХПК и общий органический углерод (ТОС) на этом первом этапе были снижены как минимум на 96%.
(2002) в промышленном масштабе после предварительных исследований в пилотном масштабе (Noronha, Britz, Mavrov, Janke, & Chmiel, 2002).Отработанные воды малых и средних предприятий смешивались в уравнительном баке и очищались с помощью пластинчатого осветлителя для удаления большей части твердых частиц. Осветленные сточные воды обрабатывали в MBR, оборудованном погруженными трубчатыми мембранами MF (размер пор 0,04 мкм), в которых полностью задерживалась активная биомасса и другие твердые частицы. Хотя уровень содержания органических веществ в отработанной воде варьировался в широких пределах (от 1800 до 6600 мг / л), ХПК и общий органический углерод (ТОС) на этом первом этапе были снижены как минимум на 96%.
Затем пермеат MF был подвергнут двухэтапному NF-процессу с интегрированной ультрафиолетовой (УФ) дезинфекцией. На первом этапе NF использовались спирально-навитые мембранные модули, характеризующиеся низким уровнем отвода хлорида натрия (в среднем 55%), что обеспечивало снижение растворенных органических веществ до 97%. Вторую стадию фильтрации проводили с использованием мембранных модулей NF со степенью удаления хлорида натрия более 90%. Эти мембраны работали как обессоливающие мембраны с низким индексом загрязнения, снижая соленость отработанного щелока более чем на 75%.Конечный ХПК обработанной воды был ниже предела обнаружения, а ТОС не превышал предельного значения 4 мг / л. Химические и микробиологические параметры конечного пермеата соответствовали стандартам качества воды Закона о питьевой воде Германии. Предлагаемый процесс показан на рисунке 18.1. Для установки производительностью 5 м 3 / час общая стоимость очистки питьевой воды из отработанной технической воды оценивается примерно в 2,5–3,0 евро / м 3 .
Вторую стадию фильтрации проводили с использованием мембранных модулей NF со степенью удаления хлорида натрия более 90%. Эти мембраны работали как обессоливающие мембраны с низким индексом загрязнения, снижая соленость отработанного щелока более чем на 75%.Конечный ХПК обработанной воды был ниже предела обнаружения, а ТОС не превышал предельного значения 4 мг / л. Химические и микробиологические параметры конечного пермеата соответствовали стандартам качества воды Закона о питьевой воде Германии. Предлагаемый процесс показан на рисунке 18.1. Для установки производительностью 5 м 3 / час общая стоимость очистки питьевой воды из отработанной технической воды оценивается примерно в 2,5–3,0 евро / м 3 .
Рисунок 18.1. Блок-схема интегрированного мембранного процесса рециркуляции отработанной технической воды в пищевой промышленности и производстве напитков.
По материалам Noronha et al. (2002). Присутствие масел и жиров в отработанных технологических водах, образующихся при производстве маргарина, создает несколько проблем, когда эти воды подвергаются биологической очистке (например, высокие затраты на аэрацию и удаление осадка, флотацию и нанесение покрытия на очистные сооружения, а также омыление жиров в уравнительный бак). Обработка этих стоков с помощью 0.Керамические мембраны MF толщиной 2 мкм снижают начальную ХПК на 5000–10 000 мг / л ниже 250 мг / л. Полученный пермеат можно было смешать с водой с низким и средним уровнем загрязнения и затем подвергнуть биологической очистке. Концентрированный продукт был извлечен из ретентата MF и повторно использован для производства мыла после обезжиривания нефти (Chmiel, Kaschek, Blöcher, Noronha, & Mavrov, 2002).
Обработка этих стоков с помощью 0.Керамические мембраны MF толщиной 2 мкм снижают начальную ХПК на 5000–10 000 мг / л ниже 250 мг / л. Полученный пермеат можно было смешать с водой с низким и средним уровнем загрязнения и затем подвергнуть биологической очистке. Концентрированный продукт был извлечен из ретентата MF и повторно использован для производства мыла после обезжиривания нефти (Chmiel, Kaschek, Blöcher, Noronha, & Mavrov, 2002).
Процесс биологического реактора с погруженной анаэробной мембраной Kubota, разработанный в последнее десятилетие компанией Kubota Membrane Technology (Хиого, Япония), представляет собой интересное применение анаэробных MBR на предприятиях пищевой промышленности.Процесс, также описанный в главе 6, состоит из резервуара для солюбилизации и мезофильного или термофильного резервуара, оборудованного погруженными мембранами. Эти мембраны удерживают метаногенные бактерии, в то время как ингибиторы ферментации метана, такие как аммиак, могут проходить в потоке пермеата. Полученный биогаз можно использовать для нагрева воды через котлы. Пермеат и отстой можно дополнительно обрабатывать в установках аэробной очистки (Kanai, Ferre, Wakahara, Yamamoto, & Moro, 2010). Этот процесс был успешно реализован на заводах по переработке пищевых продуктов, на заводах по переработке барды для Shouchu (японский спиртной напиток из сладкого картофеля) и на предприятиях по переработке картофеля.
Полученный биогаз можно использовать для нагрева воды через котлы. Пермеат и отстой можно дополнительно обрабатывать в установках аэробной очистки (Kanai, Ferre, Wakahara, Yamamoto, & Moro, 2010). Этот процесс был успешно реализован на заводах по переработке пищевых продуктов, на заводах по переработке барды для Shouchu (японский спиртной напиток из сладкого картофеля) и на предприятиях по переработке картофеля.
Ng, Lin, Panchangam, Hong и Yang (2011) сравнили производительность нового мембранного реактора с биозахватом (BEMR), заполненного носителями биошариков, с характеристиками обычного MBR при очистке сточных вод пищевых и завод по производству напитков. Новый реактор показал более длинную SRT и более низкую MLSS по сравнению с традиционной системой. BEMR производил также меньше растворимых микробных продуктов, чем традиционная система (на 34–48% меньше белка и на 16–29% меньше углеводов) из-за медленно растущих микроорганизмов с длительным SRT.Наконец, новый BEMR смог работать при постоянном потоке пермеата, что потребовало менее частой химической очистки, с вытекающими отсюда экономическими преимуществами.
Техническая осуществимость извлечения и концентрирования винной кислоты с помощью ED из сточных вод производства фруктовых соков была оценена Андресом, Риерой и Альваресом (1997). В частности, ЭД-обработка синтетических растворов, имитирующих ионообменную регенерационную воду, полученную при переработке виноградного сока, увеличила в пять раз исходную концентрацию тартрата ионов (с 1 до 10 кг / м 3 до 53.2 кг / м 3 ). Потребление энергии составило около 5 × 10 3 кДж / кг винной кислоты. Очищенную кислоту можно повторно использовать в пищевых и фармацевтических целях.
Оптимальное расположение пластинок в жабрах рыб
Значение
Обычно предполагается, что формы, встречающиеся в природе, эволюционировали таким образом, чтобы максимизировать устойчивость вида. Тем не менее, учитывая заведомо сложный дизайн природы, часто неясно, что оптимизируется.Пластинчатый узор жабр рыб — один из немногих случаев, когда оптимизация в природе может быть четко определена. Мы демонстрируем, что пластинчатая структура жабр рыб была оптимизирована, так что рыбы демонстрируют межламеллярные пространства одинакового размера независимо от массы тела или вида, тем самым показывая первичное эволюционное давление на жабры рыб. Эта естественная стратегия оптимизации демонстрирует, как контроль расположения каналов в микрофлюидных устройствах улучшает тепломассообмен.
Мы демонстрируем, что пластинчатая структура жабр рыб была оптимизирована, так что рыбы демонстрируют межламеллярные пространства одинакового размера независимо от массы тела или вида, тем самым показывая первичное эволюционное давление на жабры рыб. Эта естественная стратегия оптимизации демонстрирует, как контроль расположения каналов в микрофлюидных устройствах улучшает тепломассообмен.
Abstract
Рыбы дышат через жабры, которые в процессе эволюции извлекают водный кислород.Жабры рыб состоят из волокон с хорошо упорядоченной пластинчатой структурой, которые играют роль в максимальной диффузии кислорода. Интересно, что, когда мы анатомически наблюдаем жабры различных видов рыб, межламеллярные расстояния между жабрами ( d ) мало различаются между ними, несмотря на большие различия в массе тела ( M b ). Отмечая, что небольшие каналы, образованные плотно упакованными ламелями, вызывают значительное вязкое сопротивление потоку воды, мы построили и протестировали модель скорости переноса кислорода в зависимости от размеров ламелей и давления нагнетания, что позволяет нам предсказать оптимальное межламеллярное расстояние, которое максимизирует скорость переноса кислорода в жабрах. Сравнение нашей теории с биологическими данными подтверждает гипотезу о том, что жабры рыб эволюционировали, чтобы сформировать оптимальные межламеллярные расстояния для максимального переноса кислорода. Это объясняет слабую масштабную зависимость d от M b : d ∼ M b 1/6 .
Сравнение нашей теории с биологическими данными подтверждает гипотезу о том, что жабры рыб эволюционировали, чтобы сформировать оптимальные межламеллярные расстояния для максимального переноса кислорода. Это объясняет слабую масштабную зависимость d от M b : d ∼ M b 1/6 .
Жабры рыб эволюционировали исключительно у водных существ для извлечения кислорода из воды. Поскольку кислород имеет значительно низкую растворимость и коэффициент диффузии в воде, эффективность дыхания имеет решающее значение (1).Жабры состоят из пластинчатых структур, называемых нитями, которые покрыты массивом ламелей, окружающих сеть капиллярной крови, как показано на рис. 1 (1, 2). Богатая кислородом вода проходит через узкие каналы, образованные пластинчатыми слоями, по которым кислород диффундирует в капилляры. Плотно упакованная пластинчатая структура выгодна тем, что обеспечивает большую площадь поверхности для переноса кислорода; однако он также создает значительное вязкое сопротивление.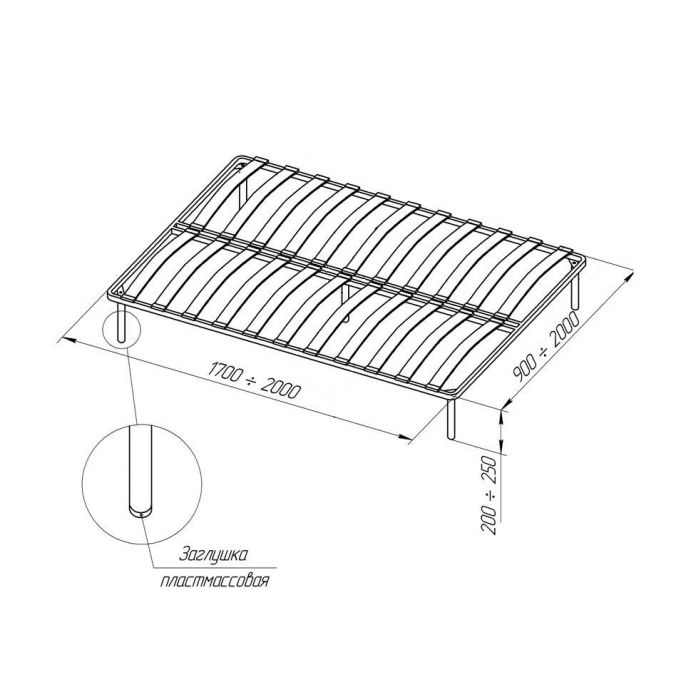 Это сопротивление преодолевается перекачкой.Рыбы обычно используют один из следующих двух насосных механизмов: жаберное откачивание и вытяжная вентиляция. Большинство костистых рыб, принадлежащих к разнообразной группе рыб с лучевыми плавниками, используют жаберное откачивание, а сжатие мускулов в глотке позволяет воде течь через жабры. При вентиляции барана, которую используют многие пелагические рыбы, динамическое давление, создаваемое их плаванием, заставляет поток воды течь в жабры (3).
Это сопротивление преодолевается перекачкой.Рыбы обычно используют один из следующих двух насосных механизмов: жаберное откачивание и вытяжная вентиляция. Большинство костистых рыб, принадлежащих к разнообразной группе рыб с лучевыми плавниками, используют жаберное откачивание, а сжатие мускулов в глотке позволяет воде течь через жабры. При вентиляции барана, которую используют многие пелагические рыбы, динамическое давление, создаваемое их плаванием, заставляет поток воды течь в жабры (3).
Жаберный аппарат морского окуня Sebastes schlegelii .( A ) Sebastes schlegelii с жабрами по обеим сторонам глотки. ( B ) Жаберная дуга и волокна. ( C ) СЭМ-изображение жаберной нити. ( D ) Поперечное сечение жаберной нити, окрашенное H и E. ( E ) Схематическое изображение морфологии жабр. Пластинчатые нити, свисающие на жаберных дугах, покрыты пластинками, окружающими сеть кровеносных капилляров. Синие стрелки указывают направление потока воды от жаберных дуг к жаберной крышке. Хорошо упорядоченные пластинчатые структуры образуют массивы микроканалов, по которым кислород диффундирует к капиллярам. ( F ) Схематическое изображение вида сверху межламеллярных каналов. Пунктирная рамка соответствует контрольному объему для анализа массопереноса.
Хорошо упорядоченные пластинчатые структуры образуют массивы микроканалов, по которым кислород диффундирует к капиллярам. ( F ) Схематическое изображение вида сверху межламеллярных каналов. Пунктирная рамка соответствует контрольному объему для анализа массопереноса.
Большинство предыдущих исследований строения жабр рыб было сосредоточено на зависимости общей площади поверхности жабр от размера тела и вида рыб (1, 4, 5). Мы рассматриваем конвективный перенос кислорода, происходящий в жабрах рыб.Когда вода проходит через узкие пластинчатые каналы, повышенное вязкое сопротивление препятствует потоку воды при заданном давлении нагнетания, которое ограничивается силой мышц или скоростью плавания; это приводит к снижению скорости переноса кислорода. Следовательно, скорость потока в промежутках ламелей и увеличенная площадь поверхности играют важную роль в определении скорости переноса кислорода. Количество ламелей на единицу длины жаберной нити определяет как площадь поверхности для диффузии, так и размер водных каналов. Поэтому мы исследуем взаимосвязь между ламеллярным расстоянием и скоростью переноса кислорода, аспект, который ранее редко исследовался.
Поэтому мы исследуем взаимосвязь между ламеллярным расстоянием и скоростью переноса кислорода, аспект, который ранее редко исследовался.
Результаты
Теоретический анализ.
Мы составили межламеллярные расстояния для широкого диапазона видов рыб, как показано на рис. 2. Примечательно, что в то время как масса тела этих видов колеблется на шесть порядков от 0,1 г до 100 кг, межламеллярные расстояния варьируются в пределах очень маленький диапазон ∼20–100 мкм (6, 7).Чтобы объяснить относительно однородные межламеллярные расстояния, мы математически смоделировали скорость переноса кислорода в жабрах рыб, которая определяется градиентом парциального давления кислорода. Для бесконечно малого контрольного объема, показанного на рис.1 F , сохранение кислорода можно записать как dQ o / dx = — hßs ( P w — P b ), где Q o — расход кислорода, h — коэффициент конвективного массопереноса, β — коэффициент растворимости кислорода в воде, s — смоченный периметр воды. контрольный объем, P w среднее парциальное давление кислорода в воде и P b парциальное давление кислорода на пластинчатой поверхности.Расход кислорода можно выразить через расход воды через канал Q w как Q o = Q w βP w . Поскольку высота ламелей H ∼400 мкм обычно намного больше, чем межламеллярное расстояние d ∼40 мкм, как показано на рис. 2 (2, 8⇓⇓ – 11), s ∼2 H , что позволяет записать QwdPw / dx + 2hH (Pw-Pb) = 0.[1]
контрольный объем, P w среднее парциальное давление кислорода в воде и P b парциальное давление кислорода на пластинчатой поверхности.Расход кислорода можно выразить через расход воды через канал Q w как Q o = Q w βP w . Поскольку высота ламелей H ∼400 мкм обычно намного больше, чем межламеллярное расстояние d ∼40 мкм, как показано на рис. 2 (2, 8⇓⇓ – 11), s ∼2 H , что позволяет записать QwdPw / dx + 2hH (Pw-Pb) = 0.[1]
Распределение межламеллярных расстояний для 75 видов рыб. Расстояние между жабрами у большинства видов рыб составляет от 20 до 110 мкм, несмотря на огромные различия в массе их тела, от 10 -1 до 10 5 г. Мы предположили, что толщина ламелей пренебрежимо мала по сравнению с межламеллярным расстоянием, и оценили межламеллярное расстояние d как обратное число ламелей на единицу длины жаберной нити (2, 6).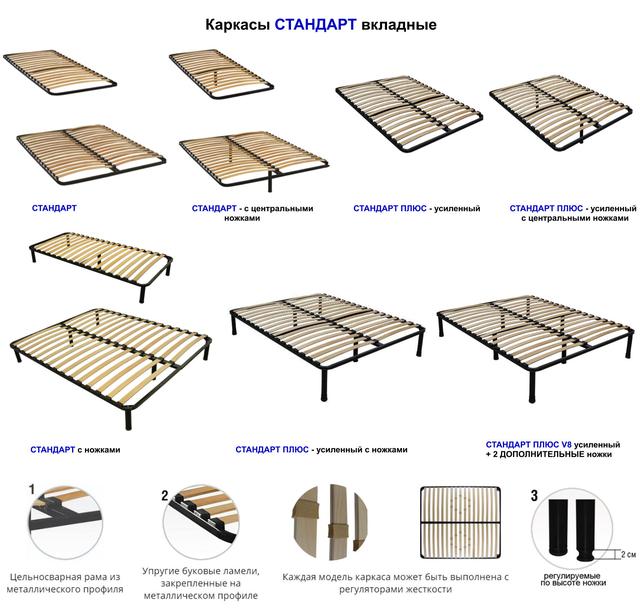 Подробные данные и их источники представлены в Таблице S1.
Подробные данные и их источники представлены в Таблице S1.
Для характерной скорости потока через межламеллярные каналы u ∼0,01 м / с (2, 11), плотность воды ρ ∼1000 кг / м 3 , вязкость μ ∼0.001 Па · с, межламеллярное расстояние d ∼10 мкм, а длина ламелей l ∼1 мм, отношение инерционных эффектов к вязким, заданное числом Рейнольдса Re = ρud 2 / ( мкл ) ∼10 — 3 , означает, что поток внутри межламеллярного канала является ламинарным с незначительными входными эффектами.Соответственно, h равно h = ShD w /2 d , где Sh — число Шервуда, отношение конвективного массопереноса к диффузионному (12), которое оценивается. Так как 7,5 и D w , коэффициент диффузии кислорода в воде (13) составляет 2 × 10 −9 м 2 / с.
Для решения уравнения. 1 для P w как функция x , сначала рассмотрим зависимость x от P b .Ожидается, что P b приблизительно соответствует парциальному давлению кислорода в капиллярах из-за незначительного диффузионного сопротивления между поверхностью ламелей и капиллярами (1). Гемоглобин в капиллярах быстро соединяется с молекулами кислорода, стабилизируя парциальное давление кислорода на относительно низком уровне (14-17). Следовательно, относительное изменение парциального давления в капиллярах относительно водяного Δ P b / Δ P w << 1, что позволяет пренебречь изменением P b с x .Решение уравнения. 1 дает P w ( x ) = ( P wo — P b ) exp ( — 2 bhx / Q w ) + P b , где P wo — парциальное давление кислорода на входе x = 0.
1 для P w как функция x , сначала рассмотрим зависимость x от P b .Ожидается, что P b приблизительно соответствует парциальному давлению кислорода в капиллярах из-за незначительного диффузионного сопротивления между поверхностью ламелей и капиллярами (1). Гемоглобин в капиллярах быстро соединяется с молекулами кислорода, стабилизируя парциальное давление кислорода на относительно низком уровне (14-17). Следовательно, относительное изменение парциального давления в капиллярах относительно водяного Δ P b / Δ P w << 1, что позволяет пренебречь изменением P b с x .Решение уравнения. 1 дает P w ( x ) = ( P wo — P b ) exp ( — 2 bhx / Q w ) + P b , где P wo — парциальное давление кислорода на входе x = 0. Поскольку скорость переноса кислорода в кровеносные капилляры в одиночном межламеллярном канале Q w β ( P wo — P w ( l )), общая скорость переноса кислорода в жабрах нить, имеющая межламеллярные каналы N , определяется как M o ∼ Q w β ( P wo — P w 9002 4 ( l )) Н. Хотя толщина ламелей t может зависеть от вида рыб или зрелости особи, мы предполагаем, что t / d <1 и N ∼ L / ( d + t ) ∼ L / d , о чем свидетельствуют многие виды, такие как Thunnus albacares , Micropterus dolomieu и Tetrapturus audax (2, 6).
Поскольку скорость переноса кислорода в кровеносные капилляры в одиночном межламеллярном канале Q w β ( P wo — P w ( l )), общая скорость переноса кислорода в жабрах нить, имеющая межламеллярные каналы N , определяется как M o ∼ Q w β ( P wo — P w 9002 4 ( l )) Н. Хотя толщина ламелей t может зависеть от вида рыб или зрелости особи, мы предполагаем, что t / d <1 и N ∼ L / ( d + t ) ∼ L / d , о чем свидетельствуют многие виды, такие как Thunnus albacares , Micropterus dolomieu и Tetrapturus audax (2, 6).
Сохранение импульса в потоке может быть описано решением потока Пуазейля, Q w = Hd 3 Δ P h / (12 мкл ), где Δ P h — перепад давления в канале (рис. 1 E ). Объединив это решение с вышеупомянутыми выражениями для M o и P w , мы наконец получили M o как функцию от d и других параметров, включая μ , D w , l и Δ P h . Дифференциация M o относительно d , таким образом, дает оптимальные межламеллярные расстояния d , которые максимизируют M o : d4 = 72µDwl2 / ΔPh.[2] Поскольку μ и D w являются свойствами материала, наша модель предполагает, что оптимальное расположение ламелей зависит исключительно от длины ламелей l и давления нагнетания Δ P h .
1 E ). Объединив это решение с вышеупомянутыми выражениями для M o и P w , мы наконец получили M o как функцию от d и других параметров, включая μ , D w , l и Δ P h . Дифференциация M o относительно d , таким образом, дает оптимальные межламеллярные расстояния d , которые максимизируют M o : d4 = 72µDwl2 / ΔPh.[2] Поскольку μ и D w являются свойствами материала, наша модель предполагает, что оптимальное расположение ламелей зависит исключительно от длины ламелей l и давления нагнетания Δ P h .
Экспериментальная проверка.
Мы проверили справедливость нашей теории для M o как функции d и Δ P h , проведя эксперименты с использованием микрожидкостного чипа, имитирующего жабры рыбы. Как показано на рис. 3 A , инженерная жабра была построена из нескольких слоев полидиметилсилоксана (PDMS), так что богатый кислородом водный канал был зажат между двумя соседними каналами противотока обедненной кислородом воды. Кислород диффундировал через тонкие мембраны PDMS от богатых кислородом до обедненных кислородом водных потоков, которые закачивались в каналы с помощью шприцевых насосов. Измерение концентрации кислорода на выходе из богатого кислородом канала для различных размеров канала и давления нагнетания дало скорость переноса кислорода M o в зависимости от d и Δ P h для фиксированной длины нити L = 1 мм.Результаты, показанные на рис. 3 B , показывают, что эмпирические измерения подтверждают нашу теорию.
Как показано на рис. 3 A , инженерная жабра была построена из нескольких слоев полидиметилсилоксана (PDMS), так что богатый кислородом водный канал был зажат между двумя соседними каналами противотока обедненной кислородом воды. Кислород диффундировал через тонкие мембраны PDMS от богатых кислородом до обедненных кислородом водных потоков, которые закачивались в каналы с помощью шприцевых насосов. Измерение концентрации кислорода на выходе из богатого кислородом канала для различных размеров канала и давления нагнетания дало скорость переноса кислорода M o в зависимости от d и Δ P h для фиксированной длины нити L = 1 мм.Результаты, показанные на рис. 3 B , показывают, что эмпирические измерения подтверждают нашу теорию.
Измерение скорости переноса кислорода с помощью микрожидкостного чипа, имитирующего жабры рыбы. ( A ) Схематическое изображение микроканалов PDMS. Богатая кислородом вода проходит через центральный канал, зажатый двумя водными каналами, обедненными кислородом. Кислород диффундирует через мембраны PDMS. ( B ) Скорость переноса кислорода M o vs.межламеллярное расстояние d для различных давлений откачки. Линии и кружки соответствуют теоретическим предсказаниям и экспериментальным результатам соответственно. Числа Рейнольдса варьируются от 0,002 до 0,3. Показаны характерные полосы погрешностей.
Богатая кислородом вода проходит через центральный канал, зажатый двумя водными каналами, обедненными кислородом. Кислород диффундирует через мембраны PDMS. ( B ) Скорость переноса кислорода M o vs.межламеллярное расстояние d для различных давлений откачки. Линии и кружки соответствуют теоретическим предсказаниям и экспериментальным результатам соответственно. Числа Рейнольдса варьируются от 0,002 до 0,3. Показаны характерные полосы погрешностей.
Обсуждение
Наш результат моделирования, уравнение. 2 , позволяет коррелировать расстояние между ламелями и длину при заданном давлении нагнетания. Давление перекачки в жабрах рыб в первую очередь зависит от насосных механизмов.Сообщается, что для большинства костистых рыб, использующих жаберное откачивание, давление откачки составляет от 5 до 50 Па (18). Пелагические костистые рыбы, которые обычно используют вытяжную вентиляцию, плавают с крейсерской скоростью v = 0,7–2 м / с, что приводит к динамическому давлению (1/2) ρv 2 ∼0,2–2 кПа, что превышает жаберное давление.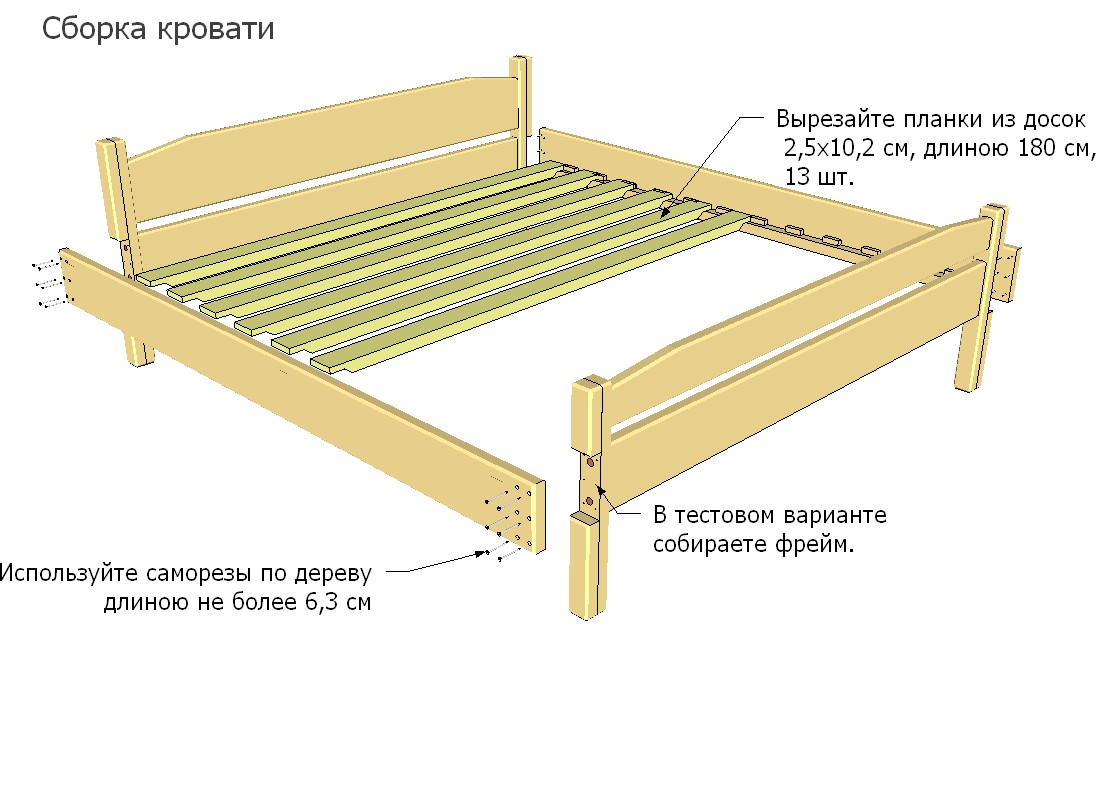 давление (19). Среди видов, использующих таранную вентиляцию, у некоторых пелагических двухжаберных рыб развилась уникальная жаберная структура, называемая межжаберной перегородкой; эта структура создает первичное сопротивление потоку воды, так что только небольшая часть динамического давления порядка 20 Па прикладывается к межламеллярным каналам (11).
давление (19). Среди видов, использующих таранную вентиляцию, у некоторых пелагических двухжаберных рыб развилась уникальная жаберная структура, называемая межжаберной перегородкой; эта структура создает первичное сопротивление потоку воды, так что только небольшая часть динамического давления порядка 20 Па прикладывается к межламеллярным каналам (11).
Мы сравнили предложенную модель Ур. 2 , с биологическими данными (рис. 4). Сравнение межламеллярного расстояния и длины пластинок в жабрах 75 видов показано на рис. 4 A . Большинство точек данных сгруппированы в пределах ожидаемого диапазона межламеллярных расстояний для каждого механизма накачки, что согласуется с нашей теорией. На рис. 4 B показана корреляция между межламеллярным расстоянием и массой тела. Предполагая геометрическое сходство между видами рыб (20), характерная длина тела должна быть пропорциональна кубическому корню из массы тела M b .Мы обнаружили, что длина ламелей l действительно масштабируется как M b 1/3 , несмотря на различия в форме жабр (1) (рис. 4 B ). На основании уравнения. 2 , который предсказывает, что оптимальное межламеллярное расстояние d должно быть масштабировано как l 1/2 для данного условия давления, мы получаем d ∼ M b 1/6 . Это масштабирование, очень слабая зависимость d от M b , объясняет биологические данные, представленные на рис.4 B , и, таким образом, раскрывает происхождение относительно однородного межламеллярного расстояния, превышающего шесть порядков массы тела.
4 B ). На основании уравнения. 2 , который предсказывает, что оптимальное межламеллярное расстояние d должно быть масштабировано как l 1/2 для данного условия давления, мы получаем d ∼ M b 1/6 . Это масштабирование, очень слабая зависимость d от M b , объясняет биологические данные, представленные на рис.4 B , и, таким образом, раскрывает происхождение относительно однородного межламеллярного расстояния, превышающего шесть порядков массы тела.
Биологические данные ламеллярных размеров жабр рыб. ( A ) Зависимость межламеллярного расстояния от длины пластинки. Пунктирными линиями обозначен диапазон оптимальных условий в зависимости от давления нагнетания. Давление откачки для рыб с использованием жаберного откачивания и для пелагических костистых рыб с использованием таранной вентиляции обычно составляет от 5 до 50 Па (18) и от 0. От 2 до 2 кПа (19) соответственно. Для пелагических пластиножаберных рыб, использующих таранную вентиляцию, но имеющих межжаберные перегородки, прилагаемое давление для межламеллярного канала составляет ~ 20 Па (11). ( B ) Зависимость длины ламелей и межламеллярного расстояния от массы тела. Из-за геометрического сходства видов рыб длина ламелей l составляет l ∼ M b 1/3 ; комбинируя масштабирование с предсказанием нашей модели, d ∼ l 1/2 , получаем d ∼ M b 1/6 .
От 2 до 2 кПа (19) соответственно. Для пелагических пластиножаберных рыб, использующих таранную вентиляцию, но имеющих межжаберные перегородки, прилагаемое давление для межламеллярного канала составляет ~ 20 Па (11). ( B ) Зависимость длины ламелей и межламеллярного расстояния от массы тела. Из-за геометрического сходства видов рыб длина ламелей l составляет l ∼ M b 1/3 ; комбинируя масштабирование с предсказанием нашей модели, d ∼ l 1/2 , получаем d ∼ M b 1/6 .
Наш анализ массопереноса, эксперименты с микрофлюидным чипом и сравнение с биологическими данными позволили нам определить первичное эволюционное давление в жабрах: максимизацию скорости переноса кислорода при заданном давлении откачки. Оптимум достигается на межламеллярном расстоянии, которое увеличивает площадь поверхности для диффузии кислорода, но не сильно препятствует потоку воды. Морфология рыбьих жабр иллюстрирует стратегию естественного дизайна, согласно которой изменение размера происходит за счет увеличения обменной поверхности, а не за счет изменения базового размера и геометрии обменной единицы.Подобные примеры можно найти в других биологических системах, таких как размер капилляров крови (20), размер эритроцитов (20) и хоаноцитарная система известковых губок (21). Эта естественная стратегия оптимизации, обнаруженная у рыб, демонстрирует, как контроль расположения каналов в микрожидкостных устройствах улучшает тепло- и массообмен, что является важной проблемой во многих практических ситуациях, включая микротеплообменники (22) и системы «лаборатория-на-чипе» для доставки лекарств. (23) и биохимический анализ (24).
Материалы и методы
Эксперименты по переносу массы с использованием искусственной жабры.
Мы изготовили инженерную систему жабр, состоящую из микроканалов PDMS, где перенос газа происходит через мембраны, разделяющие каналы. Структура PDMS была построена с использованием смеси 10: 1 Sylgard-184 (Dow Corning), отвержденной путем запекания в течение 30 минут при 80 ° C в вакуумной печи. Мы покрыли внутреннюю стенку верхнего и нижнего слоев 0,6 мас.% Teflon-AF (DuPont 601S2 и 3M Fluorinert Electronic Liquid FC40), чтобы предотвратить диффузию кислорода из каналов.Все три канала имели одинаковую ширину 2 мм и длину 70 мм. Толщина мембраны составляла 20 мкм. Мы изменили высоту центрального канала с 70 до 190 мкм, а высоту остальных каналов зафиксировали на уровне 200 мкм. Сульфит натрия использовался для контроля концентрации растворенного кислорода в воде. Канальный поток был достигнут с помощью шприцевого насоса (Harvard PHD 22/2000). Числа Рейнольдса находятся в диапазоне от 0,002 до 0,3, а течение является ламинарным. Микросенсор кислорода (Unisense OX-100) использовался для измерения парциального давления кислорода на выходе из центрального канала.
Сканирующая электронная микроскопия.
Жабры морского окуня, Sebastes schlegelii , фиксировали в фиксирующем растворе Карновского, а затем фиксировали 1% тетроксидом осмия в 0,05 М какодилатном буфере. Образец сушили в сушильном устройстве (Baltzer CPD030) после частичной дегидратации с использованием серии градуированных этанолов. Высушенный образец был покрыт тонким слоем платины в устройстве для нанесения покрытия распылением (Bal-Tec SCD005) и исследован с использованием автоэмиссионного сканирующего электронного микроскопа (Carl Zeiss SUPRA 55VP).
Благодарности
Мы благодарим Дж. У. М. Буша за ценные обсуждения и Дж. Кима за помощь с изображениями H&E. Работа выполнена при поддержке Национального исследовательского фонда Кореи, гранты 2009-0083510, 2013034978 и 2013055323.
Сноски
Вклад авторов: W.K. и Х.-Й.К. спланированное исследование; К.П. проведенное исследование; К.П., У.К. и Х.-Й.К. проанализированные данные; and K.P., W.K., and H.-Y.K. написал газету.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/lookup/suppl/doi:10.1073/pnas.1403621111/-/DCSupplemental.
Свободно доступен в Интернете через опцию открытого доступа PNAS.
Трубные отстойники против пластинчатых отстойников
Автор статьи: Брентвуд, 19.05.2020 | Категория: Вода и сточные водыПроцесс осаждения при очистке воды и сточных вод — это физический процесс, который основан на силе тяжести для осаждения и удаления взвешенных твердых частиц.В усовершенствованных процессах осаждения твердых частиц обычно используются ламели, обычно называемые трубчатыми отстойниками или пластинчатыми отстойниками, для уменьшения вертикального расстояния, на которое частицы твердых частиц должны падать до тех пор, пока не достигнут «осаждающейся поверхности». Более высокие скорости потока, лучшее качество сточных вод и более компактные отстойники могут быть достигнуты с обоими типами оборудования для осаждения ламелей. Хотя обе работают на одних и тех же принципах осаждения твердых частиц и во многих случаях могут использоваться в качестве основы для проектирования, между двумя технологиями есть несколько заметных различий.
Характеристики трубных и пластинчатых отстойников
Трубные отстойникииспользуют несколько смежных трубчатых каналов, которые наклонены под номинальным углом 60 ° и объединяются для образования увеличенной эффективной площади отстаивания. Размер и форма трубчатых каналов зависят от производителя. Конструкция трубчатого отстойника Brentwood имеет трапециевидную форму, полученную путем термоформования листов ПВХ. Вы можете узнать больше о нашем процессе проектирования, прочитав статью Thermofformed vs.Блог компании Extruded Tube Settlers.
Напротив, пластинчатые отстойники используют серию наклонных пластин, обычно изготовленных из стали, которые расположены на расстоянии от двух до трех дюймов друг от друга под углом от 55 ° до 60 ° для образования увеличенной эффективной площади отстаивания. В результате этих конструктивных различий типичные конструктивные параметры для этих двух типов различаются.
Сравнение проектных параметров
Важно отметить, что существует разница в скорости внесения (расход / площадь покрытия, выраженная в галлонах в минуту / фут²) между трубчатыми и пластинчатыми отстойниками в результате использования разных условий эффективной площади осаждения, несмотря на схожие принципы осаждения для две технологии.Чтобы обеспечить более прямое сравнение, нормы внесения отстойников для трубок могут быть преобразованы в эквивалентную норму внесения «тарелок» с учетом предполагаемой площади поверхности труб:
Как показано в приведенной выше таблице, эквивалентные максимальные «тарелочные» нормы внесения трубчатых отстойников Brentwood находятся в том же диапазоне, что и обычные тарелочные отстойники, средняя норма внесения которых составляет 0,30. Трубчатые отстойники IFR6041 наиболее сопоставимы с пластинчатыми отстойниками по площади покрытия, поскольку длина отстойников аналогична.Однако другие модули трубчатых отстойников Brentwood способны обрабатывать требуемый поток столь же эффективно с соответствующей зоной охвата. Трубные отстойники также устраняют перекрестные и вихревые токи, что делает гидравлику потока более стабильной и эффективной.
Помимо различных подходов к проектированию, существуют другие преимущества и недостатки, которые следует учитывать при сравнении трубчатых отстойников или пластинчатых отстойников для вашего приложения:
Трубные отстойники
Преимущества:
- Легкий ПВХ обеспечивает простую установку и не требует тяжелого подъемного оборудования
- Снижение капитальных затрат за счет строительных материалов
- Применяется в неглубоких резервуарах со скоростью нижнего потока
- Срок службы от двадцати до двадцати пяти лет при надлежащем техническом обслуживании
- По окончании срока службы заменяются только отстойники из трубок из ПВХ; обычно опорные конструкции и желоба для сточных вод могут оставаться
- Отстойники для трубок достаточно сильны, чтобы по ним можно было ходить для надлежащего обслуживания
- Широкие возможности настройки для различных конфигураций бассейнов
Недостатки:
- Максимальная вертикальная высота модуля составляет 41 дюйм, что дает максимальную норму внесения 3.5 галлонов в минуту / фут²
- ПВХ должен быть защищен от чрезмерного воздействия ультрафиолета. Трубчатые отстойники Брентвуда содержат ингибиторы УФ-излучения. Защитная поверхностная решетка AccuGrid также доступна для дополнительной защиты от ультрафиолета.
Пластинчатые отстойники
Преимущества:
- Меньше ограничений по глубине
- Увеличенный срок службы за счет материала конструкции
Недостатки:
- В два-три раза выше капитальные затраты
- Более высокая стоимость установки из-за необходимости тяжелого подъемного оборудования
- Требования к более глубокому бассейну для учета скоростей нижнего потока
- Риск коррозии в определенных условиях (некоторые промышленные сточные воды)
Выбор
При выборе оборудования для осаждения ламелей необходимо принимать кумулятивное решение, основанное на производительности и долгосрочном экономическом воздействии.Трубчатые отстойники могут не соответствовать сроку службы пластинчатых отстойников, но их более низкая начальная стоимость и простота установки могут быть привлекательными для завода любого размера. Их легкие и простые в обслуживании характеристики, наряду с превосходными конструктивными возможностями по сравнению с другими вариантами трубчатых отстойников на рынке, делают оборудование Brentwood очевидным выбором.
Когда дело доходит до сбора осадка, обязательно ознакомьтесь с нашими сборщиками осадка Polychem и системой земснаряда SedVac.
Исследование расстояний между передней частью носовой ости и нософронтальным клювом, базальной пластинкой, основанием черепа и клиновидной пазухой на компьютерной томографии околоносовых пазух
Задача: Целью этого исследования было изучить расстояние между передней остью носа и базальной пластинкой (BL), основанием черепа, нософронтальным клювом (NfB) и передней стенкой клиновидной пазухи на изображениях компьютерной томографии (КТ).
Методы: Всего 160 пациентов, которые были осмотрены в отоларингологической клинике нашего учреждения, были обследованы с помощью программного обеспечения для обработки изображений отделения радиологии. Случаи, в которых костные структуры не могли быть четко видны из-за предыдущей операции или травмы, были исключены. Расстояние между передними отделами носовой ости; и NfB, ближайшая точка основания черепа во фронтальной нише (SB1), ближайшая точка BL, пересечение линии, по которой было измерено расстояние до BL, с основанием черепа (SB2) и ближайшей точкой передней стенки клиновидной пазухи (Sp) измеряли с помощью мультипланарной реконструкции.
Полученные результаты: Были исследованы изображения 140 пациентов. NfB, SB1, BL, SB2 и Sp составляли 5,14 ± 0,46 (4,15-6,38) см, 5,70 ± 0,48 (4,31-7,03) см, 4,84 ± 0,44 (3,86-5,98) см, 6,23 ± 0,50 (5,13-7,35) см. и 6,14 ± 0,46 (5,04-7,36) см соответственно.
Заключение: В нашей исследовательской группе расстояния, некоторые из которых были указаны как постоянные значения в литературе, оказались в диапазоне до 2 см.Следовательно, если расстояния между структурами должны использоваться для ориентации во время эндоскопической хирургии носовых пазух, их лучше измерять перед операцией на изображениях компьютерной томографии околоносовых пазух, а не использовать заранее определенные постоянные значения.
раскрывая роль структурной неоднородности в многомасштабной артериальной механике
J R Soc Interface. 2018 окт; 15 (147): 20180492.
Xunjie Yu
1 Кафедра машиностроения, Бостонский университет, Бостон, Массачусетс, США
Raphaël Turcotte
3 Кафедра фармакологии Оксфордского университета, Оксфорд, Великобритания
Франческа Сета
4 Секция сосудистой биологии, Школа медицины Бостонского университета, Бостон, Массачусетс, США
Яньхан Чжан
1 Кафедра машиностроения, Бостонский университет, Бостон, Массачусетс, США
2 Департамент биомедицинской инженерии, Бостонский университет, Бостон, Массачусетс, США
1 Департамент машиностроения, Бостонский университет, Бостон, Массачусетс, США
2 Департамент биомедицинской инженерии, Бостонский университет, Бостон, Массачусетс, США
3 Кафедра фармакологии Оксфордского университета, Оксфорд, Великобритания
4 Секция сосудистой биологии Школы Бостонского университета Медицина, Бостон, Массачусетс, США
Поступила в редакцию 29 июня 2018 г .; Принята в печать 20 сентября 2018 г.
Издано Королевским обществом. Все права защищены.
Эта статья цитируется в других статьях в PMC.Abstract
Микроструктурная деформация эластичных ламелей играет важную роль в поддержании гомеостаза артериальной ткани и регуляции судьбы гладкомышечных клеток сосудов. Наше исследование раскрывает микроструктурное происхождение, которое позволяет эластичным пластинчатым слоям равномерно распределять напряжения через артериальную стенку, вызванные внутрипросветным растягивающим давлением, что является фундаментальным требованием для функционирования тканей и клеток.Новый экспериментальный подход был разработан для количественной оценки пространственной организации и развертывания эластичных ламеллярных слоев при повышении давления в сонных артериях мышей путем сочетания физиологического расширения-раздувания и многофотонной визуализации. Окружное растяжение на уровне ткани было получено из анализа деформации толстостенного цилиндра. Наши результаты показывают, что раскрытие и расширение ламеллярных слоев одновременно вносят вклад в деформацию на тканевом уровне. Внутренние пластинчатые слои более волнистые и развернуты больше, чем внешние.Этот градиент волнистости компенсирует большее растяжение ткани по окружности, испытываемое на внутренней поверхности, тем самым выравнивая протяженность ламеллярного слоя через стенку артерии. Открытия этого исследования показывают важность структурной неоднородности в поддержании гомеостаза тканей через артериальную стенку и могут иметь серьезные последствия для ремоделирования сосудов при старении и заболеваниях, а также в тканевой инженерии функциональных кровеносных сосудов.
Ключевые слова: упругие ламели, микромеханика, структурная неоднородность, гомеостаз, конститутивное моделирование, многофотонная визуализация
1.Введение
Эластичное волокно представляет собой компонент внеклеточного матрикса (ВКМ), который наделяет многие соединительные ткани позвоночных уникальными механическими и биологическими функциями [1]. В артериях эластические волокна образуют концентрические слои эластичных ламелей, которые в течение жизни подвергаются миллиардам циклов растяжения. Чередующиеся слои гладкомышечных клеток прикрепляются с обеих сторон к соседним пластинчатым слоям через эластиновые отростки, которые образуют сократительно-эластичную пластинчатую единицу [2,3].Хорошо задокументировано, что организованные эластичные пластинчатые единицы критически важны для гладкомышечных клеток сосудов (VSMCs), чтобы поддерживать их состояние покоя и сократимости [4–16]. Нарушение эластических волокон приводит к дедифференцировке, миграции, пролиферации и закупорке артерий VSMCs [17].
Пластинчатые элементы поддерживают и равномерно распределяют напряжения в стенке артерии, вызванные внутрипросветным растягивающим давлением, и играют важную роль в поддержании гомеостаза тканей [1]. Распределение напряжения стенки может изменять локальную проницаемость и градиент давления, которые определяют многие важные физиологические события [18].Ранние исследования показали, что количество слоев ламелей аорты линейно пропорционально диаметру артерии: несколько слоев у мышей и более 50 слоев у человека [19]. Согласно закону Лапласа, тангенциальное натяжение стенки является произведением диаметра артерии и давления растяжения. Учитывая, что вариации среднего кровяного давления у видов довольно малы по сравнению с диапазоном диаметров стенок, среднее натяжение в каждом слое ламелей было предложено примерно одинаковым, независимо от вида [19].Эти более ранние проницательные открытия указали на важную роль эластичных ламелл как фундаментальной функциональной единицы в биологии, однако до сих пор неясно, как высокоупорядоченные концентрические ламеллярные слои в стенке артерии могут равномерно распределять напряжение в радиальном направлении через стенку артерии. . Анализ напряжений однородной толстой цилиндрической стенки, подверженной внутрипросветному давлению, показал, что внутренняя поверхность просвета испытывает более высокое напряжение / деформацию стенки, чем внешняя поверхность артериальной стенки [20].Результаты 1960-х годов свидетельствуют о существовании остаточного напряжения в артерии даже при отсутствии растягивающего давления [21], хотя происхождение остаточного напряжения до сих пор не совсем понятно. С тех пор остаточное напряжение учитывалось во многих вычислительных моделях, включая конфигурацию без напряжений, которая вызывает отрицательный градиент напряжения в стенке артерии с отрицательным напряжением на поверхности просвета [18]. С такой феноменологической компенсацией может быть достигнуто более равномерно распределенное напряжение стенки, однако на сегодняшний день нет понимания того, существуют ли какие-либо структурно встроенные механизмы, позволяющие эластичным пластинчатым слоям в артериях адаптироваться к пульсирующему кровотоку и равномерно трансмурально распределять напряжение стены.
Невозможно переоценить роль ЕСМ в поддержании сосудистого гомеостаза. Волнистость эластичных ламелей была замечена на поперечном гистологическом срезе артериальной ткани в ранних исследованиях [1]. Несколько предыдущих исследований структуры эластичных ламелей, основанных на гистологическом анализе биологической ткани, показали, что эластичные ламеллярные слои разматываются при деформации [22,23]. Однако фиксация ткани, гистологический препарат и ретракция ткани при снятии механической нагрузки могут изменить структуру внеклеточного матрикса [22,24,25].Более того, трехмерная архитектура эластичных ламелей не может быть захвачена отдельными гистологическими срезами. Микромеханика эластичных ламелей при физиологической нагрузке до конца не изучена [26], и на сегодняшний день локальная деформация на ламеллярном уровне не измерялась напрямую.
Здесь мы изучили микромеханику слоев эластичных ламелей с помощью интегрированной трехмерной многофотонной визуализации, определения механических характеристик на уровне тканей и конститутивного моделирования. Мультифотонная микроскопия использовалась для визуализации микроструктурных деформаций эластичных ламелл в сонных артериях мышей при физиологическом давлении и осевом растяжении.Трехмерная структура упругих ламелей была реконструирована с целью анализа ее пространственного распределения и развертывания при двухосном механическом нагружении. В сочетании со структурно мотивированной конститутивной моделью, которая рассматривает толстостенную цилиндрическую деформацию, наше исследование исследует структурную основу, лежащую в основе микромеханики эластичных ламелей, и ее взаимосвязь с механикой сосудов на тканевом уровне. Объединив деформацию эластичных ламелей и конститутивное моделирование, мы разработали новый подход к количественной оценке локальной деформации эластичных ламелей и обнаружили важность структурной неоднородности в многомасштабной механике ткани.
2. Методы
2.1. Подготовка образца
Все экспериментальные процедуры на животных были одобрены институциональным комитетом по уходу и использованию животных (IACUC) в Медицинском кампусе Бостонского университета. Сонные артерии осторожно выделяли у восьминедельных мышей C57BL / 6 J и очищали от жира и соединительной ткани. Артерии поддерживали при 4 ° C в 1 × фосфатно-солевом буфере (PBS), визуализировали и тестировали в течение 24 часов после сбора. От артерий отделили девять сегментов сонной артерии диаметром 5 мм.Четыре сегмента использовались для визуализации, а пять образцов использовались для механических испытаний.
2.2. Многофотонная микроскопия
Ti: сапфировый лазер с синхронизацией мод (Maitai-HP, длина волны возбуждения 800 нм, Spectra-Physics, Санта-Клара, Калифорния) использовался для генерации второй гармоники из коллагена на длине волны 400 нм (417/80 нм). и двухфотонно-возбужденная флуоресценция эластина (525/45 нм), как мы описали ранее [27]. Мощность лазера была установлена на 80 мВт, чтобы изображение было как можно глубже, не вызывая насыщения двухфотонно-возбужденной флуоресценции эластина на внешней эластичной ламелле.
Изготовлена на заказ камера для растяжения-надувания тканей, которая позволяет производить двухосную деформацию растяжения-сжатия при визуализации артерии ( a , b ). Вкратце, каждый артериальный сегмент канюлировали и закрепляли на канюлях из нержавеющей стали с помощью нити 6-O и погружали в ванну с раствором PBS при комнатной температуре во время визуализации. Артериальный сегмент подвергали воздействию PBS через впускную трубку. Давление в артерии измеряли манометром, подключенным к выходному отверстию канюли.К сегменту прикладывали продольное растяжение через микрометр, подключенный к канюле. Образцы визуализировали при внутрипросветном давлении от 0 до 120 мм рт. Ст. (0, 15, 30, 45, 60, 90 и 120 мм рт. Ст.) При осевом растяжении от 1 до 1,8 (1,0, 1,3, 1,4, 1,6 и 1,8). Каждый образец был визуализирован с полем зрения 360 × 360 мкм на глубину около 50 мкм с расстоянием 2 мкм от внешней поверхности артериальной стенки с использованием иммерсионного объектива (60 ×, NA 1.0 W, LUMPlanN, Olympus ). Полученные изображения сохранялись в стеках Z ( c — f ) и использовались для последующего анализа изображений.
Мультифотонная визуализация при механической нагрузке артерии. ( a ) Схема экспериментальной установки. Многофотонный микроскоп использовался для изображения эластичных пластинчатых слоев в сонной артерии мыши, в то время как артерия подвергалась двухосному растяжению-раздуванию. ( b ) Схема сонной артерии со слоями интимы, медиа и адвентиции. Горизонтальные срезы представляют плоскости многофотонного изображения, показанные на ( c — f ). ( c — f ) Репрезентативные многофотонные изображения сонной артерии.Серый, ГВ из коллажа; зеленый — свечение эластина; масштабная линейка: 100 мкм. ( c ) Волнистые волокна коллагена в адвентициальном слое. ( d — f ) Видны адвентициальный коллаген и медиальный эластин. Продольно волнистые эластичные ламели выделены в ( e , f ). ( г ) Схема эластичной ламели, волнистой как в продольном, так и в продольном направлениях. Зеленые волнистые линии ( g ) представляют собой срезы эластичных ламелей, появляющихся на многофотонных изображениях.
2.3. Трехмерная реконструкция эластичных ламелей и анализ изображений
Для количественной оценки раскрытия ламелей во время механической деформации была разработана специальная процедура обработки изображений с использованием MATLAB Image Processing Toolbox (версия R2013b, The MathWorks, Inc., Натик, Массачусетс). Чтобы создать вид поперечного сечения упругой ламели, сначала полученные изображения Z -стопки в радиальном направлении были реорганизованы в Y -стопку в продольном направлении в MATLAB ( a ).В каждом из изображений стека Y было выбрано пороговое значение интенсивности (5 по 8-битной шкале) для устранения фона. Затем был проведен анализ связности объектов с использованием функции bwconncomp для идентификации подключенных компонентов и удаления изолированных сигналов с размером менее 8 пикселей [28].
Трехмерная реконструкция эластичных ламелей и визуальный анализ. ( a ) Исходные многофотонные изображения Z -стека были повторно разрезаны в продольном направлении ( y -ось), чтобы показать поперечный разрез эластичных ламелей.Алгоритм обработки изображений был выполнен для восстановления пластинчатых слоев и состоял в основном из ( b ) выделения исходных точек и ( c ) отслеживания линий из исходных точек для идентификации трех слоев. ( d ) Репрезентативное изображение реконструированной эластичной ламели.
В интактных сонных артериях мышей наблюдали от трех до четырех слоев эластичных ламелей. Первые три слоя эластичных ламелей были использованы для трехмерной реконструкции и анализа.Четвертый слой нельзя было систематически визуализировать, и его восстановление было ненадежным из-за низкого отношения сигнал / шум. Поэтому анализ этого слоя не проводился. Поскольку слои эластичных ламелей имеют уменьшающуюся интенсивность света через артериальную стенку, простого метода пороговой сегментации недостаточно, чтобы отделить эластичные ламели. Учитывая, что ламеллярные слои имеют непрерывную структуру и очень тонкие (около 2 мкм), для восстановления трехмерных эластичных ламелей был использован метод отслеживания линий [29].Из профиля интенсивности флуоресценции в направлении Z были идентифицированы пики, соответствующие первым трем ламеллярным слоям ( b ). Соответствующие пиксели были извлечены как начальные точки для отслеживания линии. Затем в качестве следующей исходной точки был выбран один из восьми соседних пикселей с максимальным значением интенсивности света. Уже извлеченные пиксели были исключены из поиска. Алгоритм отслеживания линии завершается, когда все восемь соседних пикселей достигают нулевого значения ( c ).Используя этот метод, центральная линия отдельной эластичной ламели отслеживалась на каждом изображении стека Y и использовалась для восстановления трехмерного ламеллярного слоя ( d ).
Структурный анализ был выполнен на трехмерной реконструированной упругой пластине. На основе извлеченных центральных линий пластинчатых слоев (L1, L2 и L3) были рассчитаны соответствующие концентрические дуги вдоль артериальной стенки на основе аппроксимации методом наименьших квадратов [30]. Параметр прямолинейности, определяемый как соотношение между длиной подогнанных дуг L a и контурной длиной ламели L c , был рассчитан и обозначен как P s ( b ). .
2,1
Количественная оценка микроструктуры эластичных ламелей во время деформации ткани. ( a ) Реконструированные изображения поперечного сечения, показывающие эластичные ламеллярные слои L1, L2 и L3 (причем L1 является наиболее удаленным слоем) в стенке артерии при 0, 15, 30, 45, 60, 90 и 120 мм рт. Осевое растяжение составляло 1,6. Масштабная линейка: 50 мкм. ( b ) Схема трех эластичных пластинчатых слоев в стенке артерии. Также определяются межпластинчатое расстояние и параметр прямолинейности P s , который представляет собой отношение длины дуги L a к длине контура L c .( c ) Параметр прямолинейности как функция давления для трех пластинчатых слоев. ( d ) Разворачивание эластичных пластинчатых слоев в сонной артерии мыши при повышении давления от 0 до 120 мм рт.ст. при осевом растяжении 1,6. (Онлайн-версия в цвете.)
Расстояние между ламелями можно оценить, вычислив разницу между радиусами подогнанных дуг ( b ). В каждом образце параметр прямолинейности и межпластинчатое расстояние были вычислены в 200 Y -образных изображениях и усреднены по отображаемой продольной длине.Результаты по четырем образцам были дополнительно усреднены и представлены в этом исследовании. Параметр прямолинейности, раскрытие ламелей, растяжение ламелей и межпластинчатое расстояние были представлены как среднее значение ± стандартная ошибка среднего (SEM).
2.4. Механическое испытание
Для характеристики зависимости давления от диаметра сонных артерий мышей, 5-миллиметровые сегменты сонной артерии ( n = 5) были протестированы на миографе давления (DMT, 110P), который позволяет измерять внутрипросветное давление и осевую силу. измеряется во время тестирования.Образцы осторожно канюлировали и хранили в бане с PBS при комнатной температуре во время механических испытаний. Артерии были предварительно растянуты до 1,6 × длины ex vivo в продольном направлении, что очень похоже на осевое растяжение in vivo [31]. Образцы предварительно кондиционировали с помощью трех циклов повышения давления от 0 до 140 мм рт.После предварительной подготовки было выполнено механическое испытание путем увеличения внутрипросветного давления с 0 до 120 мм рт. Ст. С шагом 10 мм рт. Ст. За изменением внешнего диаметра следили с помощью камеры CCD, установленной на микроскопе. Поскольку изготовленная на заказ ванна-камера для растяжения тканей не была оборудована датчиками нагрузки, многофотонная визуализация и механические испытания не выполнялись последовательно на образцах, соответствующих возрасту.
2.5. Кинематика и уравнения равновесия
Трансмуральное распределение напряжения и растяжения рассчитывалось с учетом деформации толстостенного круглого цилиндра [20].Вкратце, градиент деформации от эталонной конфигурации без нагрузки ( R , Θ , Z ) до текущей конфигурации нагрузки ( r , θ , z ) в цилиндрических координатах определяется деформацией градиент:
2,2
где λ Z — (постоянное) осевое растяжение. Принимая допущение о несжимаемости, мы имеем det F = 1, так что
2.3
Интегрирующее уравнение (2.3) дает
2,4
Радиальное положение r в деформированной стенке артерии может быть отображено в безнагрузочную конфигурацию с помощью уравнения (2.4). Из уравнения равновесия в радиальном направлении трансмуральное давление можно рассчитать как [20]:
2,5
Где p i , p o представляют внутреннее и внешнее давления соответственно. t θθ и t rr — окружные и радиальные напряжения соответственно.Из уравнения равновесия в продольном направлении осевая сила F может быть определена из [20]:
2,6
, где t ZZ — продольное напряжение. Уравнения (2.5) и (2.6) вычислялись численно с использованием схемы интегрирования Гаусса [20].
2.6. Определяющее соотношение
Тензор напряжений Коши может быть получен как
2,7
, где p — множитель Лагранжа, I — тождественный тензор, Ψ — функция деформации-энергии и является правой функцией Коши – Грина. тензор деформации.
Структурно-мотивированная деформационно-энергетическая функция, учитывающая четыре семейства коллагеновых волокон, была использована для анализа механического поведения артериальной стенки [33].
2,8
В уравнении (2.8) C e — это параметр материала, подобный напряжению, связанный с эластином, и параметры материала, связанные с семейством волокон k th. В этой модели были рассмотрены одно окружное, одно осевое и два симметрично диагональных семейства волокон. I c — первый инвариант правого тензора Коши – Грина. C , λ i — растяжение в направлении i -го семейства волокон, определяемое по формуле
2.9
, где β — это угол семейства i -го по отношению к периферическому направлению артерии в ненагруженной конфигурации.
2.7. Оценка параметров
Восемь неизвестных параметров материала ( C e ,,,,,,, β ) были оценены путем минимизации разницы между экспериментально измеренными и рассчитанными значениями давления и осевой силы на основе уравнений (2.5) и (2.6) в виде следующей целевой функции [34]:
2.10
, где N = 15 — количество точек данных, а p m , F m — давление и осевое усилие из модели, p e , F e — экспериментально измеренные давление и осевое усилие. Целевая функция минимизируется с использованием метода прямого поиска Нелдера-Мида, реализованного в функции fminsearch в MATLAB.Затем параметры материала из пяти образцов были усреднены и использованы для расчета окружного растяжения в каждом положении ламелей.
3. Результаты
3.1. Трехмерное изображение и реконструкция эластичных пластинок
Эластичные пластинчатые слои в интактной сонной артерии мыши были визуализированы с помощью многофотонного микроскопа, в то время как артерия подверглась двухосному расширению-раздуванию ( a , b , см. Методы). Многофотонные изображения в c — f показывают волнистые волокна коллагена в адвентициальном слое сонной артерии мыши на различной глубине изображения.В медиальном слое не наблюдалось видимого сигнала коллагена. Сигнал от эластина представлял собой длинные волнистые линии. Эластичные волокна в среде сонных артерий мышей объединяются в серию концентрических ламелей, и их невозможно индивидуально разрешить с помощью нашего микроскопа. Таким образом, длинные волнистые линии, показанные в c — f , представляют собой поперечное сечение загнутых эластичных ламелей, как выделено зеленым цветом в g . Качество изображения эластина было достаточным для получения поперечных сечений эластичной ламели ( a — c , см. Методы), которые использовались для количественного структурного анализа. d показывает реконструированную полную трехмерную архитектуру эластичной ламели.
3.2. Неоднородность структуры и ламеллярное разворачивание
Три ламеллярных слоя были успешно реконструированы и использованы для количественной оценки структуры. a, показывает поперечный разрез трех пластинчатых слоев сонной артерии мыши при повышении давления от 0 до 120 мм рт. Ст. При физиологическом коэффициенте растяжения в осевом направлении 1,6. При низком давлении эластичные ламели представляли собой непрерывные концентрические волнистые дуги.По мере увеличения давления эластичные ламели распрямлялись. Структурный анализ был выполнен для количественной оценки отношения окружной волнистости эластичных ламелей к давлению с использованием параметра прямолинейности ( b , см. Методы). Пластинчатые слои, расположенные ближе к внутренней стенке, имеют более низкий параметр прямолинейности, поэтому они более волнистые, чем пластинчатые слои, расположенные ближе к внешней стенке ( c ). По мере увеличения давления параметр прямолинейности всех слоев увеличивался и постепенно приближался к значению 1.Раскладывание упругих пластинчатых слоев, λ f , которое представляет выпрямление упругих пластин, было рассчитано путем нормализации параметров прямолинейности на каждом уровне давления, P s ′, параметром прямолинейности при нулевом давлении. P s , т.е. Из d видно, что все пластинчатые слои разворачиваются под давлением, однако внутренние пластинчатые слои разворачиваются больше, чем внешние слои. Кроме того, разворачивание более заметно при более низком давлении и имеет тенденцию к плато, когда давление превышает 60 мм рт.
При повышении давления эластичные ламели разворачивались, и расстояние между слоями ламелей уменьшалось ( a ). Межпластинчатое расстояние между L1 и L2 и между L2 и L3 в ненагруженном состоянии составляло 6,2 ± 1,4 и 7,9 ± 1,4 мкм соответственно. Это расстояние уменьшилось примерно на 50%, до 3,2 ± 0,4 и 4,4 ± 1,1 мкм, соответственно, при 120 мм рт. Ст. ( a ). Межпластинчатое расстояние также уменьшилось при осевом растяжении с 3,6 ± 0,3 и 7,2 ± 0,7 мкм до 2,5 ± 0,3 и 4 мкм.6 ± 0,6 мкм при увеличении осевого растяжения с 1,3 до 1,8 при транслюминальном давлении 120 мм рт. Ст. ( b ). Сонная артерия искривлена при 90 мм рт. Ст., Когда осевое растяжение ниже 1,3.
Межпластинчатое расстояние при механической нагрузке. Межпластинчатое расстояние как функция давления ( a ), когда сонные артерии подвергались осевому растяжению на 1,6, и ( b ) осевого растяжения, когда давление поддерживалось на уровне 90 мм рт. d 1 и d 2 — расстояния между слоями L1 и L2 и слоями L2 и L3 соответственно.
3.3. Деформация на уровне ткани
Окружное растяжение на уровне ткани как функция трансмурального положения было получено путем моделирования артериальной стенки в виде толстостенного цилиндра [20]. Средние параметры модели (электронный дополнительный материал, таблица S1) были получены на основе подгонки данных испытания давления на диаметр пяти сонных артерий (электронный дополнительный материал, рисунок S1). Из и можно видеть, что окружное растяжение, λ t , уменьшается от внутренней к внешней поверхности артериальной стенки, т.е.е. пластинчатый слой ближе к внутренней стенке подвергался большей деформации на тканевом уровне, чем слой ближе к внешней стенке. Чтобы сравнить растяжение ткани по окружности с ламеллярным разворачиванием, λ t было получено при радиальном положении ламеллярных слоев L1, L2 и L3. Для этого предполагалось, что L1 в ненагруженном состоянии находится на расстоянии двух третей толщины стенки от поверхности просвета [20]. Таким образом, положение L2 и L3 может быть определено путем вычитания межпластинчатых расстояний d 1 и ( d 1 + d 2 ) из L1 соответственно.Окружное растяжение ткани λ t было затем построено как функция давления в положении ламеллярных слоев L1, L2 и L3 ( b ). Мы видим, что окружное растяжение ткани увеличивалось с давлением. Однако тенденция к увеличению постепенно замедлялась при давлении выше 60 мм рт. Пластинчатые слои, расположенные ближе к поверхности просвета, постоянно подвергались более высокой деформации на тканевом уровне.
Деформация тканевого уровня и ее связь с микромеханикой.( a ) Распределение трансмурального периферического растяжения в стенке артерии при моделировании при осевом растяжении 1,6 и давлении 120 мм рт. Маркеры показывают растяжение ткани по окружности в положении ламеллярных слоев L1, L2 и L3. ( b ) Окружное растяжение ткани как функция давления в положении ламеллярных слоев L1, L2 и L3. ( c ) Растяжение ламелей как функция давления, определенное по деформации на уровне ткани ( b ) и раскрытию пластинок ( d ), было эквивалентным во всех слоях.(Онлайн-версия в цвете.)
3.4. Взаимосвязь между деформацией ткани, раскрытием ламелей и растяжением пластинок
Окружное растяжение ткани ( b ) и ламеллярное развертывание ( d ) следовали аналогичным тенденциям роста с зависимостью давления и положения от положения ламелей, однако, ламеллярное развертывание было намного меньше, чем растяжение ткани по окружности, что указывает на то, что во время механической деформации слои эластичных ламелей не только подвергаются разворачиванию, но также растягиваются или растягиваются.Таким образом, мы предлагаем новую модель, описывающую деформацию упругого пластинчатого слоя. Когда артерия находится под давлением, окружное растяжение ткани в определенном положении ламели, λ t , может быть представлено отношением длины дуги эластичной ламели в деформированной конфигурации к длине дуги в исходной конфигурации. , L a , как показано в уравнении (3.1). Мы видим, что λ t можно разложить на произведение ламеллярного развертывания λ f и растяжения ламелей λ s :
3.1
где, λ s — отношение контурной длины упругой ламели в деформированной конфигурации к длине контура в исходной конфигурации L c . λ s характеризует изменение контурной длины упругой ламели из-за растяжения / удлинения, которое невозможно измерить. Зная λ t и λ f , λ s .Из c видно, что растяжение для ламеллярных слоев, λ s , одинаково для всех трех пластинчатых слоев на каждом уровне давления.
4. Обсуждение
Настоящее исследование приводит к нескольким новым взглядам на микромеханику эластичных ламелей и ее связь с механикой тканевого уровня в стенке артерии. Новый метод был разработан для реконструкции трехмерной архитектуры эластичных ламелл в сонных артериях мышей и использовался для количественной оценки развертывания эластичных ламелл во время двухосного растяжения-раздувания нагрузки.Наше исследование показывает, что разворачивание и растяжение эластичных пластинчатых слоев одновременно способствует деформации на тканевом уровне ( a , b ). Более того, более высокие ламели, разворачивающиеся во внутреннем слое ламелей, компенсируют большую деформацию, испытываемую на внутренней поверхности артериальной стенки, тем самым поддерживая более равномерно распределенное периферическое растяжение / напряжение в ламеллярных слоях через артериальную стенку ( c ). Наше исследование впервые устанавливает, что микроструктурная неоднородность играет важную роль в поддержании гомеостаза тканей и достижении механической однородности.
Роль микромеханики эластичных ламелей в механике артериальной ткани. ( a ) Схема ламеллы эластина, которая разворачивается и растягивается, когда артериальная стенка находится под давлением. Данные многофотонной визуализации позволили нам количественно оценить ламеллярное развертывание λ f , однако растяжение упругой ламели λ s невозможно измерить и было рассчитано с использованием уравнения (3.1). ( b ) Растяжение ламелей происходит в начале повышения давления и является основным фактором общей деформации ткани.( c ) Основная роль ламеллярного раскрытия заключается в обеспечении равномерного растяжения эластичных ламелей через артериальную стенку, в то время как деформация ткани не является равномерной. (Онлайн-версия в цвете.)
Многофотонная микроскопия [27,35–37] использовалась для изучения организации внеклеточного матрикса в кровеносных сосудах. Многофотонная микроскопия имеет возможность отображать архитектуру ВЦМ биологических тканей с минимальной подготовкой образцов [26] и используется для изучения структуры коллагена и эластических волокон в различных типах биологических тканей, таких как кожа, сухожилия, связки и кровеносные сосуды. [38–41].В предыдущих исследованиях распределение ориентации и рекрутирование волокон ECM изучались на основе двумерных изображений из проекции максимальной интенсивности изображений стопки [27,42,43]. Однако такой метод анализа не подходит для изучения пространственной структуры эластичных ламелей трубчатой формы в артериях мышей. Ориентация эластических волокон у более крупных животных весьма различна [27,44–47], однако невозможно получить ориентацию эластических волокон из сонных артерий мышей.Причина такой структурной разницы между крупными и мелкими животными еще предстоит понять. Чтобы понять микромеханику эластичных ламелей, необходимо количественно оценить трехмерную архитектуру эластичных ламелей при механической нагрузке. Трехмерная реконструкция показывает, что эластичные ламели в сонной артерии мыши являются волнистыми как в окружном, так и в продольном направлениях, что важно для артерии, чтобы приспособиться к многоосевой деформации ( d ).Волнистость эластичных ламелей была замечена на поперечном гистологическом срезе артериальной ткани в ранних исследованиях [1]. Важно отметить, что волнистые линии, которые появляются на многофотонных изображениях ( d — f ), не являются эластичными волокнами, как упоминалось в более ранних исследованиях [48,49], а фактически являются срезами волнистых концентрических листов эластичных ламелей. ( г ). Сложная структура эластичных ламелей и естественная цилиндрическая форма артерии требуют трехмерной реконструкции, чтобы выявить архитектуру ламелей перед выполнением структурного анализа.
Волнистость эластичных пластинчатых слоев уменьшается по мере увеличения внутрипросветного давления. Это согласуется с более ранними гистологическими наблюдениями [1,22,23]. Волнистость и пространственное распределение эластичных ламелей трансмурально изменяются с волнистыми эластичными пластинками по направлению к внутренней поверхности артериальной стенки ( c ). Пластинчатые слои ближе к внутренней стенке более волнистые, чем пластинчатые слои ближе к внешней стенке. Когда артерия находится под давлением, более волнистый внутренний пластинчатый слой также подвергается большему разворачиванию ( d ).На уровне ткани окружное растяжение уменьшается от внутренней к внешней стенке ( a ). Однако мы обнаружили, что окружное растяжение на уровне ткани ( b ) намного больше, чем растяжение при раскрытии ( d ). Действительно, автофлуоресценция эластина позволила количественно оценить развертывание эластичных ламелей, но не их растяжение. Это отличается от сигнала ГВГ от коллагена, который может сообщать об уровне растяжения через среднюю ориентацию молекул [50].
Наше исследование демонстрирует, что ламеллярное разворачивание и растяжение происходит одновременно, и оба вносят вклад в окружное растяжение на уровне ткани ( b ). Артерии испытывают большую нагрузку на внутреннюю поверхность артериальной стенки. Более высокие ламели, разворачивающиеся во внутреннем слое ламелей, компенсируют периферический градиент растяжения ткани и, таким образом, играют важную роль в поддержании более равномерно распределенного растяжения в пластинчатых слоях через стенку артерии ( c ).Это важно, поскольку растяжение / удлинение пластин напрямую связано с развитием напряжения в пластинчатых слоях. Это решительно поддерживает предположение Добрина [23] о том, что стенка артерии механически ведет себя как однородный материал, несмотря на его гистологическую гетерогенность. Эластин в стенке артерии организован в концентрические пластинчатые слои примерно одинаковой толщины по всей стенке артерии [1]. Пластинчатый блок предназначен для поддержки и равномерного распределения механической нагрузки в стенке артерии.Наше исследование проливает свет на важность структурной неоднородности в поддержании гомеостаза тканей. В нашем организме есть встроенные механизмы для более равномерного распределения нагрузки по сосудам.
Предыдущие исследования, основанные на гистологических изображениях, предположили, что деформация эластичных ламелей представляет собой двухэтапный процесс: от нуля до диастолического давления ламели разворачиваются; затем при давлениях выше диастолического ламели растягивались [1,22,23]. Однако наше исследование количественно показывает, что раскладывание / выпрямление эластичных ламелей, λ f , вносит вклад только в небольшую часть периферической деформации ткани, λ t , даже при низком давлении (рисунки b и d ).Тот факт, что λ s увеличивается от давления 0 мм рт. Растяжение и разворачивание эластичных ламелей происходит одновременно с деформацией, и оба вносят вклад в деформацию на уровне ткани λ t с самого начала ( c ). Эти данные раскрывают важную роль градиента радиальной волнистости в выравнивании локальных окружных напряжений и подготовке эластичных пластин к физиологическому растяжению ткани по окружности, во время которого растяжение пластин является основным фактором и напрямую связано с развитием локальных напряжений в пластинчатых слоях и, таким образом, , микросреда для клеток.Важно рассматривать эластичные ламели и эластические волокна как две структурные иерархии в стенке артерии. В то время как эластичные ламели кажутся волнистыми в поперечном сечении, существует множество доказательств того, что эластичные волокна внутри эластичных ламелей находятся под напряжением из-за соматического роста даже в условиях нулевой нагрузки [51–53]. Таким образом, эластичные волокна внутри эластичных ламелей могут растягиваться, в то время как эластичные ламели выглядят волнистыми. Таким образом, наше исследование предполагает, что при изучении механики артериальной стенки важно различать деформации отдельных структурных иерархий, а структурное и механическое соответствие следует тщательно проводить.
Волнистость и межпластинчатое расстояние — две основные характеристики эластичной организации ламелей. Помимо выпрямления и уменьшения межпластинчатого расстояния с увеличением внутрипросветного давления, мы также наблюдали уменьшение межпластинчатого расстояния при осевом растяжении при определенном давлении. Межпластинчатые расстояния d 1 и d 2 , измеренные при 90 мм рт. Ст. С осевым растяжением 1,6 в сонной артерии мыши, составляют 3,6 и 5,5 мкм (), что сопоставимо со значением 0.006 мм при 100 мм рт. Ст., О которых сообщили Волински и Глагов в аорте мыши [19] без осевого растяжения. Истончение артериальной стенки в основном связано с уменьшением межпластинчатого расстояния. Это согласуется с гистологическими наблюдениями Wolinsky & Glagov [1] аорты, зафиксированной на in vivo удлинении с растяжением и без него. Соколис и др. [22] сообщил об отсутствии изменений в пространственном распределении из-за осевого растяжения. Это несоответствие, возможно, связано с испытанием на одноосное растяжение, используемым Соколисом и др. .[22]. Несколько исследований показали, что испытания на двухосное растяжение могут лучше имитировать состояние деформации, возникающей в теле, и поэтому являются предпочтительным методом [54,32].
Поскольку локальное стрессовое состояние является важным параметром, который управляет ростом и ремоделированием тканей [55], были проведены многочисленные исследования с акцентом на стресс-анализ артериальной стенки. Такие исследования часто основаны на особой форме функции деформации и энергии, в которой медиальный слой артериальной стенки обычно считается однородным материалом.Прогнозируемое распределение напряжения, основанное на таком предположении, приводит к заметному градиенту напряжения в стенке артерии, особенно в медиальном слое, который содержит слои эластичных ламелей [56–60]. Чтобы уменьшить градиент напряжения, в модель было введено остаточное напряжение, в котором было введено состояние деформации, так что, когда артерия не повреждена, остаточное напряжение добавляет окружное сжимающее напряжение к внутренней стенке и окружное растягивающее напряжение к внешней стенке. . Во многих случаях было показано, что такого феноменологического лечения недостаточно для уравновешивания градиента напряжения [20,55].В нашем исследовании сообщается о новых наблюдениях, что эластичные ламели имеют радиальный градиент волнистости. Эта структурная неоднородность компенсирует неоднородную деформацию на тканевом уровне, тем самым выравнивая локальные окружные напряжения через стенку артерии. Таким образом, необходимо разработать модели, основанные на микроструктуре, и включить такую структурную информацию на ламеллярном уровне в существующие теории роста и ремоделирования в будущих исследованиях.
4.1. Ограничения
В этом исследовании для визуализации и механических испытаний использовались разные образцы.Таким образом, параметры модели были усреднены для расчета окружного растяжения в каждом положении ламелей. Это соответствует предыдущим исследованиям [61,62], в которых средние оцененные параметры материала использовались для прогнозирования поведения группы образцов. В идеальном случае визуализация и механические испытания должны выполняться на одной и той же артерии и, таким образом, иметь специфичную для образца структуру и функцию. Однако этого часто бывает трудно достичь, особенно в небольших артериях, таких как сонные артерии мышей, потому что образцы очень хрупкие и обычно не выдерживают многократных процессов канюляции.Хотя это видно на гистологических изображениях, многофотонная визуализация не может уловить сигнал от свободно расположенных тонких медиальных коллагеновых волокон в сонных артериях мышей [43,63]. Возможно, сигнал ГВГ слишком слаб для обнаружения. Что еще более важно, наше исследование охарактеризовало медиальный эластин, независимо от медиального и адвентициального коллагена.
5. Заключение
Микромеханика упругого пластинчатого слоя была исследована путем сочетания многофотонной визуализации, механических испытаний и конститутивного моделирования.Новый подход был разработан для количественной оценки трехмерной микроструктуры эластичных ламеллярных слоев при физиологической биаксиальной механической нагрузке растяжения-раздувания. Наше исследование демонстрирует, что растяжение и разворачивание эластичных ламелей происходит одновременно во время деформации и оба вносят свой вклад в окружное растяжение на уровне тканей. В то время как растяжение пластинок доминирует над деформацией ткани, градиент радиальной волнистости эластичных пластинчатых слоев играет решающую роль в выравнивании локальных окружных напряжений по всей стенке артерии.Изучая микромеханику эластичного ламеллярного слоя, мы обнаружили важность структурной неоднородности в поддержании гомеостаза тканей. Наше исследование выявило лежащее в основе структурное происхождение, которое позволяет эластичным пластинчатым слоям равномерно распределять нагрузки через артериальную стенку, что является фундаментальным требованием для функционирования тканей и клеток. Наше исследование также подчеркнуло важность принятия многомасштабной и мультимодальной точки зрения при анализе роли компонентов ECM в механике тканей.Использование сонной артерии мыши в качестве модели позволило нам получить доступ к пластинчатым слоям с помощью многофотонной микроскопии, сохранив при этом артерию нетронутой. Распространение результатов этого исследования на крупные артерии, вероятно, является многообещающим, поскольку, основываясь на Законе Лапласа, среднее напряжение на пластинчатую единицу аортальной среды является постоянным независимо от вида [19]. Открытия этого исследования, вероятно, вдохновят на разработку тканевых кровеносных сосудов, учитывающих структурную неоднородность, и на создание новых моделей артерий, основанных на микроструктуре, для лучшего понимания роста и ремоделирования тканей.Старение и болезни могут иметь огромное влияние на состав и структуру артериальной стенки. Наш метод визуализации и анализа может быть применен для изучения структурных и механических изменений в эластичных ламелях, а также для понимания роли структурной неоднородности в поддержании тканевого гомеостаза при ремоделировании артерий.
Благодарности
Авторы благодарят профессора Чарльза П. Линь за предоставленный доступ к микроскопу.
Вклад авторов
X.Ю., Р. и Ю.З. разработал исследование. X.Y. и Р. провели многофотонную съемку и разработали метод анализа изображений. X.Y. провели эксперименты по моделированию и механическим испытаниям. Ф.С. способствовал пробоподготовке. Ю.З. руководил всем исследованием. X.Y., Y.Z. и Р. написал рукопись. Все авторы обсудили результаты и прокомментировали рукопись.
Конкурирующие интересы
У авторов нет конкурирующих интересов.
Финансирование
Эта работа была поддержана Национальным институтом здравоохранения (R01HL098028 — Y.Z., R01HL136311 для F.S.) и Национального научного фонда (CMMI 1463390 и CAREER 0954825 для Y.Z.).
Список литературы
1. Волинский Х., Глагов С. 1964 г. Структурная основа статических механических свойств аортальной среды. Circ. Res. 14, 400–413. (10.1161 / 01.RES.14.5.400) [PubMed] [CrossRef] [Google Scholar] 2. Дэвис ЕС. 1993 г. Соединения гладкомышечной клетки с эластичной пластинкой в развивающейся аорте мыши. Роль в медиальной организации аорты. Лаборатория. Инвестировать. 68, 89–99. [PubMed] [Google Scholar] 3.Baldock C, et al. 2011 г. Форма тропоэластина, протеина с высокой степенью растяжимости, который контролирует эластичность тканей человека. Proc. Natl Acad. Sci. Соединенные Штаты Америки 108, 4322–4327. (10.1073 / pnas.1014280108) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 4. Курран М.Э., Аткинсон Д.Л., Юарт А.К., Моррис Калифорния, Лепперт М.Ф., Китинг М.Т. 1993 г. Ген эластина нарушен транслокацией, связанной с надклапанным стенозом аорты. Клетка 73, 159–168. (10.1016 / 0092-8674 (93) -P) [PubMed] [CrossRef] [Google Scholar] 5.Юарт А.К., Моррис К.А., Аткинсон Д., Джин В., Стернс К., Спаллоне П., Сток А.Д., Лепперт М., Китинг М.Т. 1993 г. Гемизиготность в локусе эластина при нарушении развития, синдром Вильямса. Nat. Genet. 5, 11–16. (10.1038 / ng0993-11) [PubMed] [CrossRef] [Google Scholar] 6. Юарт А.К., Джин В., Аткинсон Д., Моррис Калифорния, Китинг М.Т. 1994 г. Надклапанный стеноз аорты, связанный с делецией, нарушающей ген эластина. J. Clin. Инвестировать. 93, 1071–1077. (10.1172 / JCI117057) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 7.Ито С., Ишимару С., Уилсон С. 1997 г. Ингибирующее действие геля коллагена 1 типа, содержащего α-эластин, на пролиферацию и миграцию гладких мышц сосудов и эндотелиальных клеток. Кардиоваск. Surg. 5, 176–183. (10.1016 / S0967-2109 (97) 00004-5) [PubMed] [CrossRef] [Google Scholar] 8. Ито С, Ишимару С, Уилсон С.Е. 1998 г. Влияние коацервированного α-эластина на пролиферацию гладких мышц сосудов и эндотелиальных клеток. Ангиология 49, 289–297. (10.1177 / 0003319798047) [PubMed] [CrossRef] [Google Scholar] 9.Карник С.К., Брук Б.С., Байес-Генис А., Соренсен Л., Уайт Дж. Д., Шварц Р. С., Китинг М. Т., Ли Д. Я. 2003 г. Решающая роль передачи сигналов эластина в морфогенезе сосудов и заболеваниях. Разработка 130, 411–423. (10.1242 / dev.00223) [PubMed] [CrossRef] [Google Scholar] 10. Ли Д.Й., Толанд А.Е., Боак Б.Б., Аткинсон Д.Л., Энсинг Г.Дж., Моррис Калифорния, Китинг М.Т. 1997 г. Точечные мутации эластина вызывают обструктивное заболевание сосудов, надклапанный стеноз аорты. Гм. Мол. Genet. 6, 1021–1028. (10.1093 / hmg / 6.7.1021) [PubMed] [CrossRef] [Google Scholar] 11.Ли Д.Й., Брук Б., Дэвис Е.К., Мехам Р.П., Соренсен Л.К., Боак Б.Б., Эйхвальд Е., Китинг М.Т. 1998 г. Эластин является важным детерминантом артериального морфогенеза. Природа 393, 276–280. (10.1038 / 30522) [PubMed] [CrossRef] [Google Scholar] 12. Ли Д.Й., Фори Дж., Тейлор Д.Г., Дэвис Е.К., Бойл В.А., Мехам Р.П., Стензель П., Боак Б., Китинг М.Т. 1998 г. Новая артериальная патология у мышей и гемизиготных по эластину человека. J. Clin. Инвестировать. 102, 1783–1787. (10.1172 / JCI4487) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 13.Тассабехджи М., Меткалф К., Доннаи Д., Херст Дж., Рирдон В., Берч М., Рид А. П.. 1997 г. Эластин: структура генома и точечные мутации у пациентов с надклапанным стенозом аорты. Гм. Мол. Genet. 6, 1029–1036. (10.1093 / hmg / 6.7.1029) [PubMed] [CrossRef] [Google Scholar] 14. Урбан З. и др. 2000 г. Изолированный надклапанный стеноз аорты: функциональная гаплонедостаточность гена эластина в результате нонсенс-опосредованного распада. Гм. Genet. 106, 577–588. (10.1007 / s0043
285) [PubMed] [CrossRef] [Google Scholar] 15.Urbán Z, Riazi S, Seidl TL, Katahira J, Smoot LB, Chitayat D, Boyd CD, Hinek A. 2002 г. Связь между гаплонедостаточностью эластина и повышенной пролиферацией клеток у пациентов с надклапанным стенозом аорты и синдромом Вильямса – Бёрена. Являюсь. J. Hum. Genet. 71, 30–44. (10.1086 / 341035) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 16. Ямамото М., Ямамото К., Нумура Т. 1993 г. Коллаген типа I способствует преобразованию культивируемых клеток гладкой мускулатуры артерий кролика от сократительного до синтетического фенотипа.Exp. Cell Res. 204, 121–129. (10.1006 / excr.1993.1016) [PubMed] [CrossRef] [Google Scholar] 17. Брук Б.С., Байес-Генис А., Ли Д.Я. 2003 г. Новые взгляды на эластин и сосудистые заболевания. Тенденции Кардиоваск. Med. 13, 176–181. (10.1016 / S1050-1738 (03) 00065-3) [PubMed] [CrossRef] [Google Scholar] 18. Чыонг CJ, Фунг YC. 1986 г. Остаточное напряжение в артериях. In Frontiers in biomechanics (ред. Schmid-Schönbein GW, Woo SLY, Zweifach BW), стр. 117–129. Нью-Йорк, штат Нью-Йорк: Спрингер. [Google Scholar] 19. Волинский Х, Глагов С.1967 г. Пластинчатая единица медиального строения и функции аорты у млекопитающих. Circ. Res. 20, 99–111. (10.1161 / 01.RES.20.1.99) [PubMed] [CrossRef] [Google Scholar] 20. Holzapfel GA, Gasser TC, Ogden RW. 2000 г. Новая основа для механики артериальной стенки и сравнительное исследование моделей материалов. J. Elast. Phys. Sci. Твердые тела 61, 1–48. (10.1016 / S0022-3697 (99) 00252-8) [CrossRef] [Google Scholar] 22. Соколис Д.П., Кефалояннис Э.М., Кулукусса М., Маринос Э., Будулас Х., Караяннакос ЧП. 2006 г.Структурная основа взаимосвязи напряжения и деформации аорты при одноосном растяжении. J. Biomech. 39, 1651–1662. (10.1016 / j.jbiomech.2005.05.003) [PubMed] [CrossRef] [Google Scholar] 23. Добрин ПБ. 1999 г. Распространение пластинчатых деформаций: влияние на свойства артериальной среды. Гипертония 33, 806–810. (10.1161 / 01.HYP.33.3.806) [PubMed] [CrossRef] [Google Scholar] 24. Храпко М., ван Доммелен Дж. А., Петерс Г. В., Висманс Дж. С.. 2008 г. Влияние условий испытаний на характеристику механических свойств ткани головного мозга.J. Biomech. Англ. 130, 31003 (10,1115 / 1,2
- 6) [PubMed] [CrossRef] [Google Scholar] 25. Мартинес П.А., Бербель-Филхо В.М., Якобина Ю.П.
2013.
Может ли фиксация формалином и сохранение этанола влиять на геометрический морфометрический анализ? Рыбы как пример. Зооморфология
132, 87–93. (10.1007 / s00435-012-0176-x) [CrossRef] [Google Scholar] 27. Чоу MJ, Turcotte R, Lin CP, Zhang Y.
2014 г.
Артериальный внеклеточный матрикс: механобиологическое исследование вклада и взаимодействия эластина и коллагена.Биофиз. Дж.
106, 2684–2692. (10.1016 / j.bpj.2014.05.014) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 28. Цуда А., Филипович Н., Хабертан Р. Д., Дики Р., Мацуи Ю., Стампанони М., Шиттни Дж.
2008 г.
Конечно-элементная трехмерная реконструкция ацинуса легких, полученная с помощью синхротронной рентгеновской томографии. J. Appl. Physiol.
105, 964–976. (10.1152 / japplphysiol..2008) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 29. Влахос М., Дерматас Э.
2010 г.
Многоуровневая сегментация сосудов сетчатки с использованием отслеживания линий.Comput. Med. Графики изображений
34, 213–227. (10.1016 / j.compmedimag.2009.09.006) [PubMed] [CrossRef] [Google Scholar] 30. Хольцапфель Г.А., Зоммер Г., Ауэр М., Регитниг П., Огден Р.В.
2007 г.
Послойные трехмерные остаточные деформации аорты человека с неатеросклеротическим утолщением интимы. Анна. Биомед. Англ.
35, 530–545. (10.1007 / s10439-006-9252-z) [PubMed] [CrossRef] [Google Scholar] 31. Беллини С., Ферруцци Дж., Роккабьянка С., Ди Мартино Е.С., Хамфри Дж. Д..
2014 г.
Микроструктурно мотивированная модель механики артериальной стенки с механобиологическими последствиями.Анна. Биомед. Англ.
42, 488–502. (10.1007 / s10439-013-0928-x) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 32. Ферруцци Дж., Берси МР, Хамфри Дж. Д.
2013.
Биомеханическое фенотипирование центральных артерий в условиях здоровья и болезней: преимущества и методы для моделей на мышах. Анна. Биомед. Англ.
41, 1311–1330. (10.1007 / s10439-013-0799-1) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 33. Ван В., Янагисава Х., Глисон Р.Л.
2010 г.
Биомеханические и микроструктурные свойства общих сонных артерий мышей без фибулина-5.Анна. Биомед. Англ.
38, 3605–3617. (10.1007 / s10439-010-0114-3) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 34. Ли Ю, Найто Й, Куробе Х, Брейер К. К., Хамфри Дж. Д..
2013.
Биаксиальные механические свойства нижней полой вены у мышей C57BL / 6 и CB-17 SCID / bg. J. Biomech.
46, 2277–2282. (10.1016 / j.jbiomech.2013.06.013) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 35. Сугита С., Мацумото Т.
2016 г.
Наблюдения с помощью многофотонной микроскопии трехмерных изменений микроструктуры эластина и коллагеновых волокон во время повышения давления в среде аорты.Биомех. Модель. Механобиол.
16, 763–773. (10.1007 / s10237-016-0851-9) [PubMed] [CrossRef] [Google Scholar] 36. Кларк Т.Э., Лилли М.А., Фогл А.В., Гослайн Дж.М., Шедвик RE.
2015 г.
Механический вклад пластинчатого и межламеллярного эластина вдоль аорты мыши. J. Biomech.
48, 3608–3614. (10.1016 / j.jbiomech.2015.08.004) [PubMed] [CrossRef] [Google Scholar] 37. Зуми А., Лу Х, Кассаб Г.С., Тромберг Б.Дж.
2004 г.
Визуализация микроструктуры коронарной артерии с использованием второй гармонической и двухфотонной флуоресцентной микроскопии.Биофиз. Дж.
87, 2778–2786. (10.1529 / biophysj.104.042887) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 38. У С, Ли Х, Ян Х, Чжан Х, Ли З, Сюй С.
2011 г.
Количественный анализ морфологии коллагена в стареющей коже на основе многофотонной микроскопии. J. Biomed. Опт.
16, 40502 (10.1117 / 1.3565439) [PubMed] [CrossRef] [Google Scholar] 39. Frisch KE, Duenwald-Kuehl SE, Kobayashi H, Chamberlain CS, Lakes RS, Vanderby R.
2012 г.
Количественная оценка организации коллагена с использованием фрактальных размерностей и преобразований Фурье.Acta Histochem.
114, 140–144. (10.1016 / j.acthis.2011.03.010) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 40. Маттсон Дж. М., Тюркотт Р., Чжан Ю.
2016 г.
Гликозаминогликаны способствуют привлечению волокон внеклеточного матрикса и механике артериальной стенки. Биомех. Модель. Механобиол. 16, 213–225. (10.1007 / s10237-016-0811-4) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 41. Задрозный Л.М., Нойфельд Е.Б., Люкотт Б.М., Коннелли П.С., Ю З.-Х, Дао Л., Сюй Л.-Й, Балабан Р.С.
2014 г.
Исследование развития трехмерной макромолекулярной структуры грудной аорты мышей с помощью двухфотонной микроскопии.J. Histochem. Cytochem.
63, 8–21. (10.1369 / 0022155414559590) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 42. Чен Х, Слипченко М.Н., Лю Ю, Чжао Х, Ченг Дж-Х, Ланир Й, Кассаб Г.С.
2013.
Биаксиальная деформация коллагеновых и эластиновых волокон в коронарной адвентиции. J. Appl. Physiol.
115, 1683–1693. (10.1152 / japplphysiol.00601.2013) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 43. Ван В., Диксон Дж. Б., Глисон Р. Л..
2012 г.
Конститутивное моделирование сонных артерий мышей с использованием экспериментально измеренных микроструктурных параметров.Биофиз. Дж.
102, 2916–2925. (10.1016 / j.bpj.2012.04.035) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 44. Фаранд П., Гарон А., Планте Дж. Э..
2007 г.
Строение крупных артерий: ориентация эластина во внутренней эластической пластине аорты кролика и в эластических пластинках среды аорты. Microvasc. Res.
73, 95–99. (10.1016 / j.mvr.2006.10.005) [PubMed] [CrossRef] [Google Scholar] 45. Кох Р.Г., Цамис А., Дамор А., Вагнер В.Р., Уоткинс СК, Глисон Т.Г., Ворп Д.А.
2014 г.
Пользовательский инструмент анализа изображений на основе изображений для количественной оценки микроархитектуры эластина и коллагена в стенке аорты человека с помощью многофотонной микроскопии.J. Biomech.
47, 935–943. (10.1016 / j.jbiomech.2014.01.027) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 46. Красный В, Морин С, Магоариец Х, Аврил С.
2017 г.
Комплексное исследование послойных морфологических изменений микроструктуры сонных артерий при одноосной нагрузке. Acta Biomater.
57, 342–351. (10.1016 / j.actbio.2017.04.033) [PubMed] [CrossRef] [Google Scholar] 47. Ю Икс, Ван И, Чжан Ю.
2018.
Трансмуральные вариации в распределении ориентации эластиновых волокон в стенке артерии.J. Mech. Behav. Биомед. Матер.
77, 745–753. (10.1016 / j.jmbbm.2017.08.002) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 48. Фрай Дж. Л. и др.
2015 г.
Сиртуин-1 гладких мышц сосудов защищает от расслоения аорты при гипертензии, индуцированной ангиотензином II. Варенье. Сердце доц.
4, e002384 (10.1161 / jaha.115.002384) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 49. Киз Дж. Т., Хаскетт Д. Г., Утцингер У., Азхар М., Ванде Гест Дж. П.
2011 г.
Адаптация плоского микробиаксиального оптомеханического устройства для тубулярных биаксиальных микроструктурных и макроскопических характеристик мелких сосудистых тканей.J. Biomech. Англ.
133, 75001 (10.1115 / 1.4004495) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 50. Тюркотт Р., Маттсон Дж. М., Ву Дж. В., Чжан Ю., Линь С. П..
2016 г.
Молекулярный порядок артериального коллагена с использованием изображений генерации второй гармоники с круговой поляризацией. Биофиз. Дж.
110, 530–533. (10.1016 / j.bpj.2015.12.030) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 51. Мартин С., Гринвальд С.
1997 г.
Нарушение синтеза эластина в стенках аорты и крупных кондуитных артерий на раннем этапе развития как исходное событие в патогенезе системной гипертензии.Ланцет
350, 953–955. (10.1016 / S0140-6736 (96) 10508-0) [PubMed] [CrossRef] [Google Scholar] 53. Ферруцци Дж., Коллинз М.Дж., Йе А.Т., Хамфри Дж. Д..
2011 г.
Механическая оценка целостности эластина в сонных артериях с дефицитом фибриллина-1: последствия для синдрома Марфана. Кардиоваск. Res.
92, 287–295. (10.1093 / cvr / cvr195) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 54. Цзоу И, Чжан Ю.
2009 г.
Экспериментальное и теоретическое исследование анизотропии эластиновой сети. Анна. Биомед. Англ.
37, 1572–1583.(10.1007 / s10439-009-9724-z) [PubMed] [CrossRef] [Google Scholar] 55. Fung YC.
1991 г.
Что делают остаточные напряжения в наших кровеносных сосудах?
Анна. Биомед. Англ.
19, 237–249. (10.1007 / BF02584301) [PubMed] [CrossRef] [Google Scholar] 56. Хаяси К., Сато М., Ниими Х., Ханда Х., Моритаке К.
1975 г.
Анализ основных законов сосудистой стенки с помощью теории конечных деформаций. Иёдэнши То Сэйтай Когаку
13, 293–298. [PubMed] [Google Scholar] 57. Хаяси К.
1982 г.
Фундаментальные и прикладные исследования механических свойств сердечно-сосудистых тканей.Биореология
19, 425–436. (10.3233 / BIR-1982-19304) [PubMed] [CrossRef] [Google Scholar] 58. Чыонг CJ, Фунг YC.
1983 г.
Трехмерное распределение напряжения в артериях. J. Biomech. Англ.
105, 268 (10,1115 / 1,3138417) [PubMed] [CrossRef] [Google Scholar] 59. фон Мальцан В.В., Варрияр Р.Г., Кейтцер В.Ф.
1984 г.
Экспериментальные измерения эластических свойств сред и адвентиций сонных артерий крупного рогатого скота. J. Biomech.
17, 839–847. (10.1016 / 0021-9290 (84) -8) [PubMed] [CrossRef] [Google Scholar] 60.Такамидзава К., Хаяси К.
1987 г.
Функция плотности энергии деформации и гипотеза равномерной деформации для механики артерий. J. Biomech.
20, 7–17. (10.1016 / 0021-9290 (87) -4) [PubMed] [CrossRef] [Google Scholar] 61. Ван Ц., Гарсия М., Лу Х, Ланир И, Кассаб Г.С.
2006 г.
Трехмерные механические свойства коронарных артерий свиней: проверенная двухслойная модель. Являюсь. J. Physiol. Слушайте Circ. Physiol.
291, 1200–1209. (10.1152 / ajpheart.01323.2005) [PubMed] [CrossRef] [Google Scholar] 62. Коллинз М.Дж., Берси М., Уилсон Э., Хамфри Д.Д.2011 г.
Механические свойства надпочечников и инфраренальной брюшной аорты: значение для мышиных моделей аневризм. Med. Англ. Phys.
33, 1262–1269. (10.1016 / j.medengphy.2011.06.003) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 63. Montes GS, Nicolelis MAL, Brentani-Samaia HP, Furuie SS.
1989 г.
Диаметр коллагеновых фибрилл в артериях мышей. Клетки Ткани Органы
135, 57–61. (10.1159 / 000146723) [PubMed] [CrossRef] [Google Scholar]
- 1.
Шмитц, Г. и Мюллер, Г. Структура и функция ламеллярных телец, липидно-белковые комплексы, участвующие в хранении и секреции клеточных липидов. J. Lipid Res. 32 , 1539–1570 (1991).
CAS PubMed Статья PubMed Central Google ученый
- 2.
Дэниэлс, К. Б. и Оргейг, С. Легочный сурфактант: ключ к эволюции дыхания воздухом. N. Physiol. Sci. 18 , 151–157 (2003).
CAS Google ученый
- 3.
Фесслер, М. Б. и Саммер, Р. С. Поверхностно-активные липиды на границе раздела «хозяин-среда». метаболические сенсоры, супрессоры и эффекторы воспалительных заболеваний легких. Am. J. Respir. Cell Mol. Биол. 54 , 624–635 (2016).
CAS PubMed PubMed Central Статья Google ученый
- 4.
Эррера-Рамос, Э.и другие. Генетические варианты поверхностно-активного белка А связаны с тяжелой респираторной недостаточностью при инфекции вирусом пандемического гриппа А. Crit. Уход 18 , R127 (2014).
PubMed PubMed Central Статья Google ученый
- 5.
Glasser, S. W. et al. Мыши с дефицитом поверхностно-активного протеина С восприимчивы к респираторно-синцитиальной вирусной инфекции. Am. J. Physiol. Lung Cell Mol. Physiol. 297 , L64–72 (2009).
CAS PubMed PubMed Central Статья Google ученый
- 6.
Флорос, Дж. И Томас, Н. Дж. Генетика поверхностно-активных белков при внебольничной пневмонии: балансирование воспалительного состояния хозяина. Crit. Уход 15 , 156 (2011).
PubMed PubMed Central Статья Google ученый
- 7.
Виейра, Ф., Кунг, Дж. У.И Бхатти, Ф. Структура, генетика и функция легочных ассоциированных сурфактантных белков A и D: внелегочная роль этих лектинов С-типа. Ann. Анат. 211 , 184–201 (2017).
PubMed PubMed Central Статья Google ученый
- 8.
Лет-Ларсен, Р., Чжун, Ф., Чоу, В. Т., Холмсков, У. и Лу, Дж. Спайк-гликопротеин коронавируса SARS избирательно распознается белком D сурфактанта легких и активирует макрофаги. Иммунобиология 212 , 201–211 (2007).
CAS PubMed PubMed Central Статья Google ученый
- 9.
Woods, P. S. et al. Смертельная инфекция вируса гриппа А h2N1 изменяет липидом сурфактанта альвеолярных клеток II типа. Am. J. Physiol. Lung Cell Mol. Physiol. 311 , L1160 – L1169 (2016).
PubMed PubMed Central Статья Google ученый
- 10.
Sun, X. et al. Метаболизм жирных кислот связан с тяжестью заболевания после инфицирования H7N9. EBioMedicine 33 , 218–229 (2018).
PubMed PubMed Central Статья Google ученый
- 11.
Ochs, M. Чем ближе мы смотрим, тем больше видим? Количественный микроскопический анализ системы легочных сурфактантов. Cell Physiol. Biochem. 25 , 27–40 (2010).
CAS PubMed Статья PubMed Central Google ученый
- 12.
Grathwohl, C., Newman, G.E., Phizackerley, P.J. и Town, M.H. Структурные исследования пластинчатых осмиофильных тел, выделенных из легкого свиньи. Результаты 31P ЯМР и содержание воды. Biochim. Биофиз. Acta 552 , 509–518 (1979).
CAS PubMed Статья PubMed Central Google ученый
- 13.
Voorhout, W.F., Weaver, T.E., Haagsman, H.P., Geuze, H.J. и Van Golde, L.M. Биосинтетическая маршрутизация белков легочного сурфактанта в клетках альвеолярного типа II. Microsc. Res. Tech. 26 , 366–373 (1993).
CAS PubMed Статья PubMed Central Google ученый
- 14.
Hobi, N. et al. Маленький ключ открывает тяжелую дверь: основная функция небольших гидрофобных белков SP-B и SP-C — запускать адсорбцию легочных пластинчатых тел сурфактанта. Biochim. Биофиз. Acta 1863 , 2124–2134 (2016).
CAS PubMed Статья PubMed Central Google ученый
- 15.
Mulugeta, S. et al. Идентификация LBM180, мембранного белка, ограничивающего пластинчатое тело альвеолярных клеток типа II, в качестве белка-переносчика ABC ABCA3. J. Biol. Chem. 277 , 22147–22155 (2002).
CAS PubMed Статья PubMed Central Google ученый
- 16.
Мацумура Ю., Сакаи, Х., Сасаки, М., Бан, Н. и Инагаки, Н. Поглощение холин-фосфолипидов, опосредованное ABCA3, во внутриклеточные везикулы в клетках A549. FEBS Lett. 581 , 3139–3144 (2007).
CAS PubMed Статья PubMed Central Google ученый
- 17.
Перес-Гил, Дж. Структура легочных мембран и пленок сурфактанта: роль белков и липид-белковые взаимодействия. Biochim. Биофиз. Acta 1778 , 1676–1695 (2008).
CAS PubMed Статья PubMed Central Google ученый
- 18.
Vanhecke, D. et al. Пересмотр ультраструктуры пластинчатого тела: замораживание под высоким давлением и криоэлектронная микроскопия срезов стекловидного тела. Histochem. Cell Biol. 134 , 319–326 (2010).
CAS PubMed Статья PubMed Central Google ученый
- 19.
Аль-Амуди, А., Студер, Д. и Дубочет, Дж. Вырезание артефактов и процесс резки в срезах стекловидного тела для криоэлектронной микроскопии. Дж.Struct. Биол. 150 , 109–121 (2005).
CAS PubMed Статья PubMed Central Google ученый
- 20.
Rigort, A. et al. Микрообработка эукариотических клеток сфокусированным ионным пучком для криоэлектронной томографии. Proc. Natl Acad. Sci. США 109 , 4449–4454 (2012).
CAS PubMed Статья PubMed Central Google ученый
- 21.
Марко, М., Шей, К., Шалек, Р., Франк, Дж. И Маннелла, К. Разрежение замороженных гидратированных биологических образцов с помощью фокусированного ионного пучка для криоэлектронной микроскопии. Nat. Методы 4 , 215–217 (2007).
CAS PubMed Статья PubMed Central Google ученый
- 22.
Kukulski, W. et al. Коррелированная флуоресценция и трехмерная электронная микроскопия с высокой чувствительностью и пространственной точностью. Дж.Cell Biol. 192 , 111–119 (2011).
CAS PubMed PubMed Central Статья Google ученый
- 23.
Schorb, M. & Briggs, J. A. Коррелированная криофлуоресценция и криоэлектронная микроскопия с высокой пространственной точностью и повышенной чувствительностью. Ультрамикроскопия 143 , 24–32 (2014).
CAS PubMed Статья PubMed Central Google ученый
- 24.
Мецкас, Л. А. и Бриггс, Дж. А. Г. Обнаружение состояния слияния мембран на крио-ЭМ сетке на основе флуоресценции с использованием коррелированной криофлуоресценции и криоэлектронной микроскопии. Microsc. Микроанал. 25 , 942–949 (2019).
CAS PubMed PubMed Central Статья Google ученый
- 25.
Sartori-Rupp, A. et al. Корреляционная криоэлектронная микроскопия выявляет структуру TNT в нейрональных клетках. Nat. Commun. 10 , 342 (2019).
CAS PubMed PubMed Central Статья Google ученый
- 26.
Плитцко, Дж. М., Ригорт, А. и Лейс, А. Корреляционная крио-световая микроскопия и крио-электронная томография: от клеточных территорий до молекулярных ландшафтов. Curr. Opin. Biotechnol. 20 , 83–89 (2009).
CAS PubMed Статья PubMed Central Google ученый
- 27.
Schellenberger, P. et al. Высокоточная корреляционная флуоресцентная и электронная криомикроскопия с использованием двух независимых маркеров совмещения. Ультрамикроскопия 143 , 41–51 (2014).
CAS PubMed PubMed Central Статья Google ученый
- 28.
Hampton, C.M. et al. Коррелированная флуоресцентная микроскопия и криоэлектронная томография инфицированных вирусом или трансфицированных клеток млекопитающих. Nat.Protoc. 12 , 150–167 (2017).
CAS PubMed Статья PubMed Central Google ученый
- 29.
Arnold, J. et al. Пробоподготовка криофокусированного ионного пучка в зависимости от места проведения под контролем трехмерной корреляционной микроскопии. Biophys. J. 110 , 860–869 (2016).
CAS PubMed PubMed Central Статья Google ученый
- 30.
Wilfling, F. et al. Селективный путь аутофагии для фазовых отложений эндоцитарных белков. Мол. Клетка. 80 , 764–778.e7 (2020).
CAS PubMed PubMed Central Статья Google ученый
- 31.
Lieber, M., Smith, B., Szakal, A., Nelson-Rees, W. & Todaro, G. Непрерывная линия опухолевых клеток из карциномы легкого человека со свойствами альвеолярного эпителия типа II. клетки. Внутр.J. Cancer 17 , 62–70 (1976).
CAS PubMed Статья PubMed Central Google ученый
- 32.
Wolff, G. et al. Не забывайте о зазоре: швы с микродомпенсированием значительно уменьшают изгиб крио-ламелей, изготовленных фрезерованием FIB. J. Struct. Биол. 208 , 107389 (2019).
PubMed Статья PubMed Central Google ученый
- 33.
Castano-Diez, D., Kudryashev, M., Arheit, M. & Stahlberg, H. Dynamo: гибкий и удобный инструмент разработки для усреднения субтомограмм крио-электромагнитных данных в высокопроизводительных вычислительных средах. J. Struct. Биол. 178 , 139–151 (2012).
PubMed Статья PubMed Central Google ученый
- 34.
Cooper, J. R. et al. Долгосрочное культивирование линии раковых клеток A549 способствует формированию многослойных телец и дифференцировке по фенотипу пневмоцитов альвеолярного типа II. PLoS ONE 11 , e0164438 (2016).
PubMed PubMed Central Статья CAS Google ученый
- 35.
Gorelick, S. et al. PIE-область, интегрированная криокорреляционная световая и микроскопия FIB / SEM. Элиф https://doi.org/10.7554/eLife.45919 (2019).
- 36.
Кришнамурти, Г., Ван, К. Ю., Стейер, Г. и Уилсон, Д. Л. Удаление подповерхностной флуоресценции при крио-визуализации с использованием деконволюции. Опт. Экспресс 18 , 22324–22338 (2010).
PubMed PubMed Central Статья Google ученый
- 37.
Faoro, R. et al. Криоиммерсионная световая микроскопия с коррекцией аберраций. Proc. Natl Acad. Sci. США 115 , 1204–1209 (2018).
CAS PubMed Статья PubMed Central Google ученый
- 38.
Вольф, Г., Хаген, К., Грюневальд, К. и Кауфманн, Р. К корреляционной флуоресценции сверхвысокого разрешения и электронной криомикроскопии. Biol. Ячейка 108 , 245–258 (2016).
CAS PubMed PubMed Central Статья Google ученый
- 39.
Haller, T., Cerrada, A., Pfaller, K., Braubach, P. & Felder, E. Микроскопия в поляризованном свете выявляет физиологические и вызванные лекарствами изменения в сборке сурфактантной мембраны в пневмоцитах альвеолярного типа II. Biochim. Биофиз. Acta Biomembr. 1860 , 1152–1161 (2018).
CAS PubMed Статья PubMed Central Google ученый
- 40.
Mahamid, J. et al. Жидкокристаллические фазовые переходы в липидных каплях связаны с клеточными состояниями и специфической ассоциацией органелл. Proc. Natl Acad. Sci. США 116 , 16866–16871 (2019).
CAS PubMed Статья PubMed Central Google ученый
- 41.
Engelbrecht, S., Kaltenborn, E., Griese, M. & Kern, S. Липидный переносчик сурфактанта ABCA3 расщепляется на N-конце внутри LAMP3-положительных везикул. FEBS Lett. 584 , 4306–4312 (2010).
CAS PubMed Статья PubMed Central Google ученый
- 42.
Bhowmick, R. et al. Трехмерная тканевая модель легких человека для изучения инфекции гриппа А. Tissue Eng. Часть A 24 , 1468–1480 (2018).
CAS PubMed PubMed Central Статья Google ученый
- 43.
Zhang, L. et al. Rab38 нацеливается на ламеллярные тела и нормализует их размеры в эпителиальных клетках альвеолярного типа II легких. Am. J. Physiol. Lung Cell Mol. Physiol. 301 , L461–477 (2011).
CAS PubMed PubMed Central Статья Google ученый
- 44.
Tanaka, H. et al. Структура свода печени крысы с разрешением 3,5 ангстрем. Наука 323 , 384–388 (2009).
CAS PubMed Статья PubMed Central Google ученый
- 45.
Rast, A. et al. Биогенные участки тилакоидов цианобактерий образуют участки контакта с плазматической мембраной. Nat. Растения 5 , 436–446 (2019).
CAS PubMed Статья PubMed Central Google ученый
- 46.
Preibisch, S., Saalfeld, S. & Tomancak, P. Оптимальное в глобальном масштабе сшивание мозаичных 3D-изображений для получения микроскопических изображений. Биоинформатика 25 , 1463–1465 (2009).
CAS PubMed PubMed Central Статья Google ученый
- 47.
Schindelin, J. et al. Фиджи: платформа с открытым исходным кодом для анализа биологических изображений. Nat. Методы 9 , 676–682 (2012).
CAS Статья Google ученый
- 48.
Мастронарде, Д. Н. Автоматизированная электронно-микроскопическая томография с надежным предсказанием движений образца. J. Struct. Биол. 152 , 36–51 (2005).
Артикул Google ученый
- 49.
Хаген, У. Дж. Х., Ван, У. и Бриггс, Дж. А. Г. Реализация схемы наклона криоэлектронной томографии, оптимизированной для усреднения субтомограмм высокого разрешения. J. Struct. Биол. 197 , 191–198 (2017).
PubMed PubMed Central Статья Google ученый
- 50.
Мастронард, Д. Н. и Хелд, С. Р. Автоматическое выравнивание серии наклона и томографическая реконструкция в IMOD. J. Struct. Биол. 197 , 102–113 (2017).
PubMed Статья PubMed Central Google ученый
- 51.
Paul-Gilloteaux, P. et al. eC-CLEM: гибкое программное обеспечение для многомерной регистрации для корреляционных микроскопий. Nat. Методы 14 , 102–103 (2017).
CAS PubMed Статья PubMed Central Google ученый
v — линейный темп роста РФ,
λ α —толщина ламелей α,
N ε —количество атомов, попадающих в РФ за время τ ,
- 4498 макс. — максимальное количество атомов, попадающих в РФ,
S — количество секторов в РФ (от 0 до λ α расстояния), используемых в модели,
C p — емкость (максимальное количество атомов) каждого сектора, определенного в модели.
- 1.
v = const, N ε = const, для разных значений λ α ,
- 2.
Посткорреляционная крио-CLEM на ламелях выявляет мембранную архитектуру пластинчатых тел
О непрерывном движении прерывистого фронта осадков
Модель предполагает, что DP представляет собой комбинированную реакцию миграции RF и диффузии RF [11]. Это означает, что атомы растворенного вещества входят в границу α / α o путем диффузии вдоль RF к богатой растворенными веществами β-ламелле.Чтобы достичь равновесного состояния внутри α-ламели, все атомы растворенного вещества должны диффундировать вдоль границы на расстояние λ α /2 ( λ α — толщина α-ламели ) при перемещении РФ на собственную ширину δ с определенной скоростью v . Обычно время, отведенное для вытеснения, слишком мало, и некоторые из атомов растворенного вещества не достигают края богатых растворенными веществами β-ламелей. Они должны перепрыгнуть через RF, чтобы осуществить следующее элементарное смещение.Таким образом, атомы, которые пересекают RF, способствуют формированию профиля концентрации растворенного вещества за RF. Число атомов, которые должны диффундировать через RF, увеличивается с дальнейшим движением RF, формируя профиль растворенного вещества. По прошествии определенного времени количество атомов становится настолько большим, что это может вызвать остановку RF, чтобы позволить всем избыточным атомам войти в обедненную растворенными веществами α-ламеллу. Затем RF расслабляется, и процесс начинается снова.
Для визуализации такого процесса была подготовлена специальная процедура с использованием технологий HTML5, CSS3 и JavaScript, работающих на каждом устройстве, поддерживающем просмотр веб-сайтов (рис.2). HTML был применен как простой язык маркеров для определения позиций входных данных в браузере пользователя, в то время как CSS — это язык, используемый для описания стиля: цвета фона, размеры шрифта, стили ввода и т. Д. С другой стороны, JavaScript отвечал за для анимации процесса, например вычисления и перерисовки экрана в заданный интервал времени. Также есть два основных холста, которые используются для отображения процесса и результирующих параметров. Все приложение упаковано в один файл, что позволяет легко копировать и запускать его на разных устройствах.
Рис. 2Блок-схема процедуры моделирования
Модель может моделировать различные случаи с использованием параметров, которые могут быть установлены пользователем, например:
Следует подчеркнуть, что моделирование принимает во внимание каждый сектор и, используя приближение наименьших квадратов, можно найти функцию, которая лучше всего подходит к данным. Процесс моделирования также можно приостановить, продолжить и сбросить в любое время. В результате получают профили концентрации растворенного вещества, оставшиеся после движущейся вперед RF прерывистых осадков. Они описываются с использованием модели DP реакции Кана [13]. Основная идея этой модели — уравновешивание масс растворенных атомов, на которые влияет диффузия вдоль RF и движение RF.{2}}} {{s \ delta D _ {\ text {b}}}} $$
(2)
Здесь x o — исходное содержание растворенного вещества в сплаве, x i — содержание растворенного вещества в α-ламелле, контактирующей с β-ламелью, v — линейная скорость роста РФ, λ α — толщина α-ламели, с — фактор сегрегации, δ — ширина границы зерен (РФ), D b — диффузия по границам зерен коэффициент, а y — координата, измеренная от края β-ламели в направлении, перпендикулярном α-ламели.
За исключением параметров C , моделирование дает: τ go — время, в течение которого RF движется, τ остановка — время, необходимое для того, чтобы все избыточные RF атомы вошли в обедненную растворенными веществами α-ламеллу (иными словами время, в течение которого ВЧ неподвижен) и τ итого — время всей продолжительности процесса.
Моделирование роста прерывных выделений проводилось на примере Al-22 ат.% сплава, в котором процесс DP был подробно описан, включая наблюдение на месте [14, 15] и систематические исследования химии через ламели RF и α-фазы с использованием аналитической электронной микроскопии [9, 16]. Входные параметры для уравнения. 1 были приняты равными x o = 22 ат.% И x i = 4,4 ат.%. Были рассмотрены два случая: